Tartalom
- Fejlődési mintázatképzés (pattern formation)
- A toolkit és az állatok testfelépítése
- Áttörés a fejlődésbiológiában — A Drosophila korai fejlődésének analízise
- Áttörés az evolúciós kutatás felé - Hox-gének és a modern EVO-DEVO hajnala
- A genetikai „toolkit” - együtt a testterv kialakításáért felelős gének
- A homológia fogalmának folyamatos evolúciója
- Mit fogadunk el bizonyítéknak az EVO-DEVO-ban?
- Morfológiai evolúció és génszabályozás
- A toolkit kialakulása: az első állatok
- Közös ősünk - az Urbilateria rekonstrukciója
- A testtervek és a testrészek diverzifikációja
- A Hox fehérjék és az általuk szabályozott gének
- A négylábúak eredete
- Hox-gének: descent with modification
A toolkit-génkészletnek, ill. annak elemeinek felfedezése kétségtelenül a biológia legnagyobb horderejű és legszélesebb horizontokat nyitó, új paradigmákat hozó fejezetei közé tartozik. A történeti hűséghez hozzátartozik, hogy számos nagy felfedezés időben párhuzamosan történt, és hogy a kép természetesen fokozatosan, de utólag tekintve gyorsan és lenyűgöző hatékonysággal tisztult le.
A felfedezések döntő része nem kimondottan evolúciós szemszögből történt, de a kép teljességéhez hozzátartoznak ezek az eredmények is. A fejezetben áttekintjük azokat a fejlődésbiológiai eredményeket is, amelyek fontosak a „toolkit”-koncepció nagyszerűségének megértéséhez, vállalva, hogy a teljességre itt reményünk sincsen. Az érdeklődők a következő fogalmakról teljesebb képet alkothatnak fejlődésbiológia tárgyú könyvek tanulmányozásával.
A fejlődésbiológia célja annak a folyamatnak a megértése, melynek során a petéből felnőtt egyed alakul ki.
Mai értelmezés szerint tulajdonképpen a fejlődésnek minden tér- és időbeli valamint minőségváltozással kapcsolatos vonatkozása a fejlődési mintázatképzés (pattern formation) tárgykörbe tartozik.
Szűkebb értelemben e folyamat során alakul ki a majdan differenciálódó sejtek elrendeződése az élőlényben, előrevetítve a celluláris differenciáció térbeliségét.
A „testterv” (Bauplan, body plan) a biológia egyik legnehezebb fogalma, itt csak intuitív megközelítésére vállalkozunk:
Egyes nagyobb élőlénycsoportok – „monofiletikus taxonok” – testének részei mindig meghatározott módon rendeződnek el, függetlenül a környezettől. Ez az elrendeződés a testterv.
A testterv kialakulásának így mindenképpen örökletes alapjai vannak, tehát érdemes genetikai módszerekkel tanulmányozni. 1. ábra
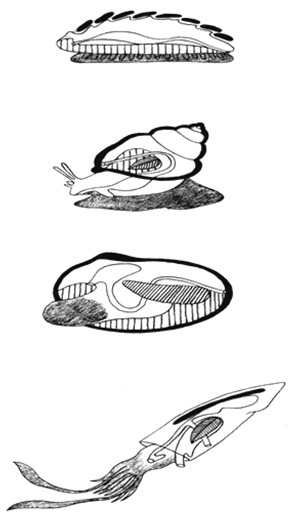
A fejlődési mintázatképzés néven összefoglalt folyamatok felelősek az összetett forma kialakulásáért, és a testterv fejlődési programjának kivitelezéséért.
A fejlődési mintázatképzés kulcsfogalma a POZICIONÁLIS INFORMÁCIÓ.
Van egy olyan sejtparaméter, a POZICIONÁLIS ÉRTÉK, amely hozzárendelhető a fejlődő rendszerben az adott sejt pozíciójához. Értelmezéséhez mindig szükség van egy viszonyítási alapra („koordináta rendszerre” vagy inkább egy határvonalra, „boundary-re”), amelyhez a sejtek pozíciójukat viszonyítják. Genetikai állományuk közvetlenül nem tartalmaz olyan információkat, amelyek lokalizációjukat „közölnék” a sejtekkel 2. ábra.
10.2. ábra - A csirkeszárny és a légy szárnya egyaránt pozicionális jelet használ mintázata kialakításakor. A pozicionális jel egy morfogén, amely az A/P tengely mentén hat. A csirkénél a jel a polarizáló régióból indul, míg a légynél a jel forrása a kompartmenthatár; a jelet a sejtek másképpen értelmezik az elülső-és hátsó kompartmentben
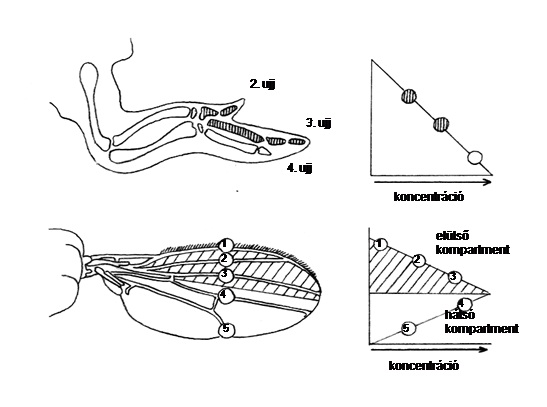 |
A pozícionális információ egyik vonzó tulajdonsága általános felhasználhatósága. Ugyanaz a koordinátarendszer és ugyanaz a szignál többször is felhasználható, akár ugyanabban az embrióban. Az emlősök elülső és hátulsó végtagjaiban és a legyek szárnyában hasonló módon molekuláris szignálok határozzák meg a pozícionális információt (az A/P azaz anterio-poszteriorális és D/V azaz dorzoventrális viszonyokat)
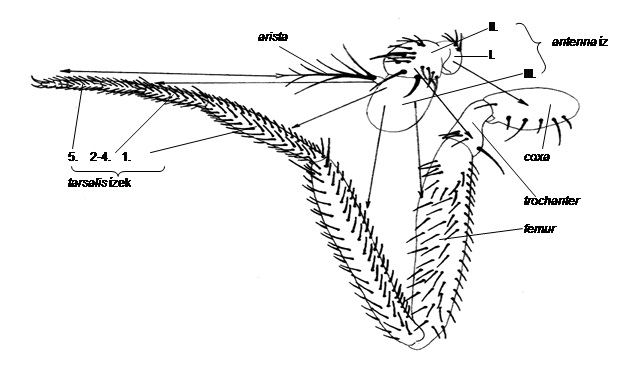
A pozicionális információnak másik alapvető sajátsága az, hogy független attól, hogy a sejtek miként differenciálódnak, vagyis nincsen rokonság a pozicionális érték és a differenciáció módja között. A sejtek tehát pozicionális információjukat fejlődési útjuknak és genotípusuknak (epigenotípusuknak) megfelelően értelmezhetik. A pozicionális értékek azonosak pl. a rovarok lábában és csápjában, mint azt a későbbiekben látni fogjuk, de interpretációjuk másként történik.3. ábra, 4. ábra
10.3. ábra - Megfelelés a csáp és a láb részei között. Az Antennapedia mutánsban a csáp bizonyos régiói lábstruktúrákká transzformálódtak. A nyilak a csáp régióit mutatják, amelyek csakis a láb nekik megfelelő struktúráivá transzformálódtak. Hasonló megfelelés volt tapasztalható a génkifejeződés szintjén is
 |
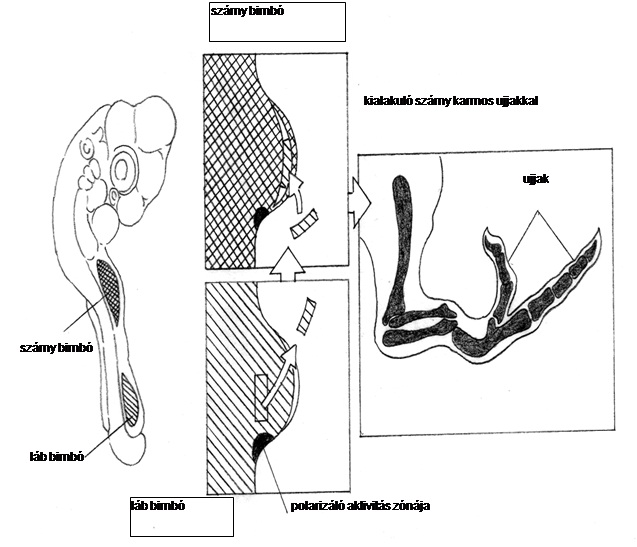
10.4. ábra - A proximális láb-bimbó sejteket a szárny-bimbó disztális részébe ültetve azok a progresszív zónában disztálisabb pozícionális értéket nyernek az eredetinél, és karmos ujjakat alakítanak ki comb helyett.
 |
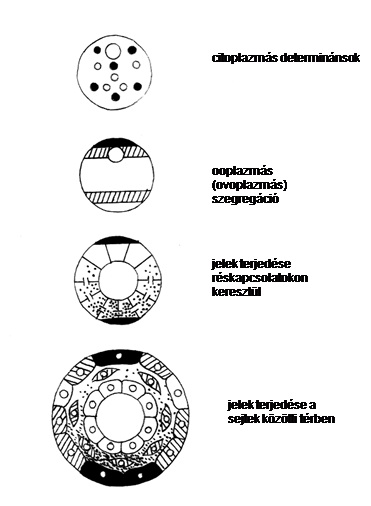
Mintázatkialakítás és determináció. Klasszikus kísérleteken alapuló hipotetikus séma. A fejlődés első szakaszában a specifikáció és elköteleződés (commitment) a citoplazmatikus determinánsok révén történik. A citoplazmatikus determinánsok térben a pete belső mintázatképzési folyamata, az ooplazmás szegregáció folyamán rendeződnek. Egy részük morfogénként működik, és a pólusokon lokalizálódik, megteremtve a morfogéngrádiens-képzés lehetőségét. A morfogének különböző koncentrációi különféle elsődleges zigótikusan aktív regulátorgéneket aktiválnak és az embriót néhány széles doménre osztják. A másodlagos szabályozógének kifejeződési doménjei az embriót felosztják még finomabb doménokra, különféle sejtsorsokkal. A sejtidentitási gének aktiválása és aktivitásuk fenntartása lehetővé teszi a sejteknek, hogy sorsukat fenntartsák fejlődésük további részében.A hasadások folyamán más-más blasztomerekbe más-más determinánsok jutnak. A későbbi fejlődés folyamán mintázatképzési-és determinációs folyamatok (amelyek nem választhatók el) a különböző sejtek közötti jelcserén alapulnak. 5. ábra
A KOMPARTMENTALIZÁCIÓ olyan határok létét jelenti az élőlényen belül, amelyeket bizonyos sejtek utódjai (klónjai) sohasem lépnek át. A határok a legtöbb esetben nem köthetők anatómiai értelemben vett határvonalakhoz. A határok kulcsjelentőségűek, amennyiben a kompartmenthatárokból kiinduló jelek alakítják ki a kompartmentek egyes sejtjeinek pozicionális értékét. A kompartmentek sejtjei kisszámú „ALAPÍTÓ” (FOUNDER) sejttől származnak. A sejtklónok mozgásának korlátozódása az elemzett esetekben sejtjeik felszínének sajátságaira vezethetők vissza.