A homológia összehasonlító biológiai tanulmányok és szisztematika szempontjából egyaránt alapvető filogenetikai koncepciója szerint azok a homológ sajátságok, amelyek, többé vagy kevésbé módosult formában öröklődnek tovább attól a közös őstől, ahol a jelleg először kialakult. Más megközelítésben a homológ struktúrák szünapomorfiák.
A homológiára utal a más testrészekhez viszonyított helyzet, hasonlóság legalábbis néhány strukturális sajátságban, és átmeneti képletek jelenléte akár (kihalt és élő) fajok között, akár az egyedfejlődés során, ill. a felsoroltak kombinációja. A leírtak megfigyelése után a homológia hipotézisét kiértékelik, annak alapján, hogy mennyire vág egybe az adott jellegből levont a más jellegek alapján kikövetkeztetett leszármazással.
Így az várható, hogy a genetikai és fejlődési szempontból hasonló jellegek homológok, és a filogenetikai szempontból homológ jellegeknek hasonlóak a fejlődési és genetikai alapjai. Meglepő módon sok megfigyelés ellentétben áll ezekkel a feltevésekkel, ami oda vezetett, hogy további koncepciót vezettek be, a biológiai homológia koncepcióját.
A szériaszerűen homológ (serially homologous) struktúrák, mint az ízeltlábúak szelvényei, lábai, vagy a gerincesek fogai nagyrészt ugyanazt a fejlődésgenetikai hátteret („masinériát”) használják az egyedfejlődés folyamán, de nyilvánvalóan nem „történetileg” homológok. Ennél sokkal mélyebb probléma merül fel az evolúció „visszafordulásánál” (evolutionary reversal). Látványos példa a második őrlőfog (molar) esete hiúzban. A második őrlőfog jelen van a legtöbb húsevőben, de egy macskafélében, a hiúzban eltűnt, majd újra megjelent (igaz, csak az egyedek tizedében). A hiúz második őrlőfoga nem homológ filogenetikai értelemben a többi húsevő őrlőfogával, hanem homopláziás jelleg. Fejlődésgenetikai szempontból azonban „ugyanaz” a struktúra, mint a többi húsevőé, mert a fogat létrehozó mechanizmusok nagyon hasonlóak, sőt, ugyanazok lehetnek, mint a más húsevőkben hatók.
A filogenetikai és biológiai homológiák között további konfliktust okoz, hogy a filogenetikailag homológ jellegeknek különböző fejlődési és genetikai alapjai lehetnek. A szalamandrák ujjai fordított sorrendben differenciálódnak, mint bármely más négylábúé.
Minden állati szemlencse tartalmaz különböző crystallin fehérjéket, de a különböző állatvonalak crystallinjai különböző fehérjékből alakultak ki. A fordított helyzet is előfordul: különböző taxonok fejlődési és funkcionális szempontból hasonló struktúrái filogenetikailag előfordulhat, hogy nem homológok. A legklasszikusabb példa, hogy az állati szemek egymástól függetlenül fejlődtek ki a legkülönbözőbb állatoknál, amit régen egyértelműen analógiaként tárgyaltak (azonos funkció, független evolúciós fejlődés). Bonyolítja a képet, hogy minden egyes taxonban a rendkívül konzervatív Pax6 játszik alapvető szerepet a szem fejlődésében. Később ennek jelentőségére visszatérünk.
A biológiai homológia koncepciója arra épül, hogy egy sajátság a szerveződés bizonyos, például fenotípus szinten homológ a fajok között, míg más szinten, például fejlődési vagy genetikai szinten nem beszélhetünk homológiáról. A koncepció azt az alapelvet, hogy a soksejtű élőlények többé-kevésbé konzervált eszközök (az angol a „tool” szót használja) készletéből állnak össze. Az „eszközök” lehetnek egyedi gének, fehérjék, vagy sokféle fehérjéből álló szabályozókörök („circuit”). A morfológiai evolúció nagy részben, ahogy azt a Nobel-díjas Jacob a hetvenes években oly szellemesen megfogalmazta: „barkácsolás” („tinkering”) ezzel az eszközkészlettel (ld. Makroevolúció). Az EVO-DEVO egyik legnagyobb kihívása annak megértése, hogyan fejlődnek ki a biológiailag homológ strukturális egységek az eszközök adott készletéből 21. ábra.
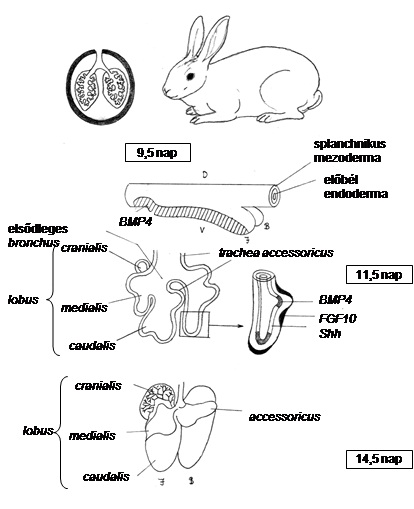
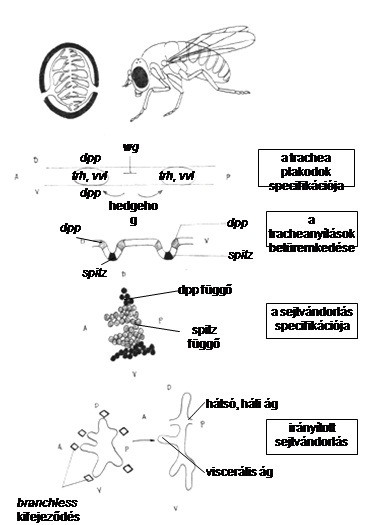
10.21. ábra - A tüdő fejlődésének genetikai és funkcionális alapjai evolúciósan nagymértékben konzerváltak. A Drosophila és az emlősök légzőszervének fejlődésében oly fontos epitélium-kommunikáció részletbe menően hasonlóan zajlik. A Drosophila modellnek bizonyult egy eddig gerinces-specifikusnak tartott szerv tanulmányozásához. A Drosophilákkal közös őseink öröksége minden korábbi elképzelést meghaladóan meghatározza fejlődésünket és létünket. Az emlősök tüdejének esetében az FGF10, a BMP4 és a Sonic hedgehog gének kifejeződési mintázata a tüdő kialakulása során azok kritikus szerepe mellett szól
 |
 |