A különböző élőlények fejlődési génjeinek és genomjának analízise megmutatta, hogy már az egér és Drosophila közös ősének, azaz az ős- és újszájúak közös ősének is komoly toolkitje volt fejlődési génekből. A homeodoménnel rendelkező gének jelen vannak az összes többsejtűben, bár funkciójuk korántsem ugyanaz, pl. a növényekben nem látnak el az állatokéihoz hasonló globális funkciókat.
Vannak olyan teljes jelátviteli útvonalak, amelyek csak állatokban fordulnak elő pl. a TGF-β, Wnt szignálmolekulák.
A metazoák legközelebbi „outgroupjai” a Choanoflagellaták (soha ne becsüljük le az egysejtűeket: van közöttük, amelynek kétszázszor akkora genomja van, a Tetrahymenanának pedig kb. ugyanannyi génje – 27000 - mint nekünk), „bazális” állati taxonok a csalánozók (Cnidaria), és a még távolabbiak, a szivacsok (Porifera).
A galléros ostorosok (Choanoflagellaták) egysejtűek, vannak jelátviteli útvonalaik, adhéziós molekuláik, amelyek már a toolkit kialakulásának kezdeteit jelzik.
További toolkit gének jelennek meg a Cnidariaknal: TGF-β, Wnt szignálmolekulák, transzkripciós faktor családok, homeodomén gének, amelyek homológok a bilateráliákéihoz (even-skipped, engrailed, Hox, 4 Pax, T-boksz (Brachyury), snail, twist).
-
A CNIDARIÁK ÉS BILATERÁLÁK TOOLKITJÉNEK ÖSSZEVETÉSE MEGMUTATTA, HOGY A TOOLKIT KITERJEDT A CNIDÁRIÁK ELKÜLÖNÜLÉSE UTÁN, DE A BILATERÁLIÁK RADIÁCIÓJA ELŐTT
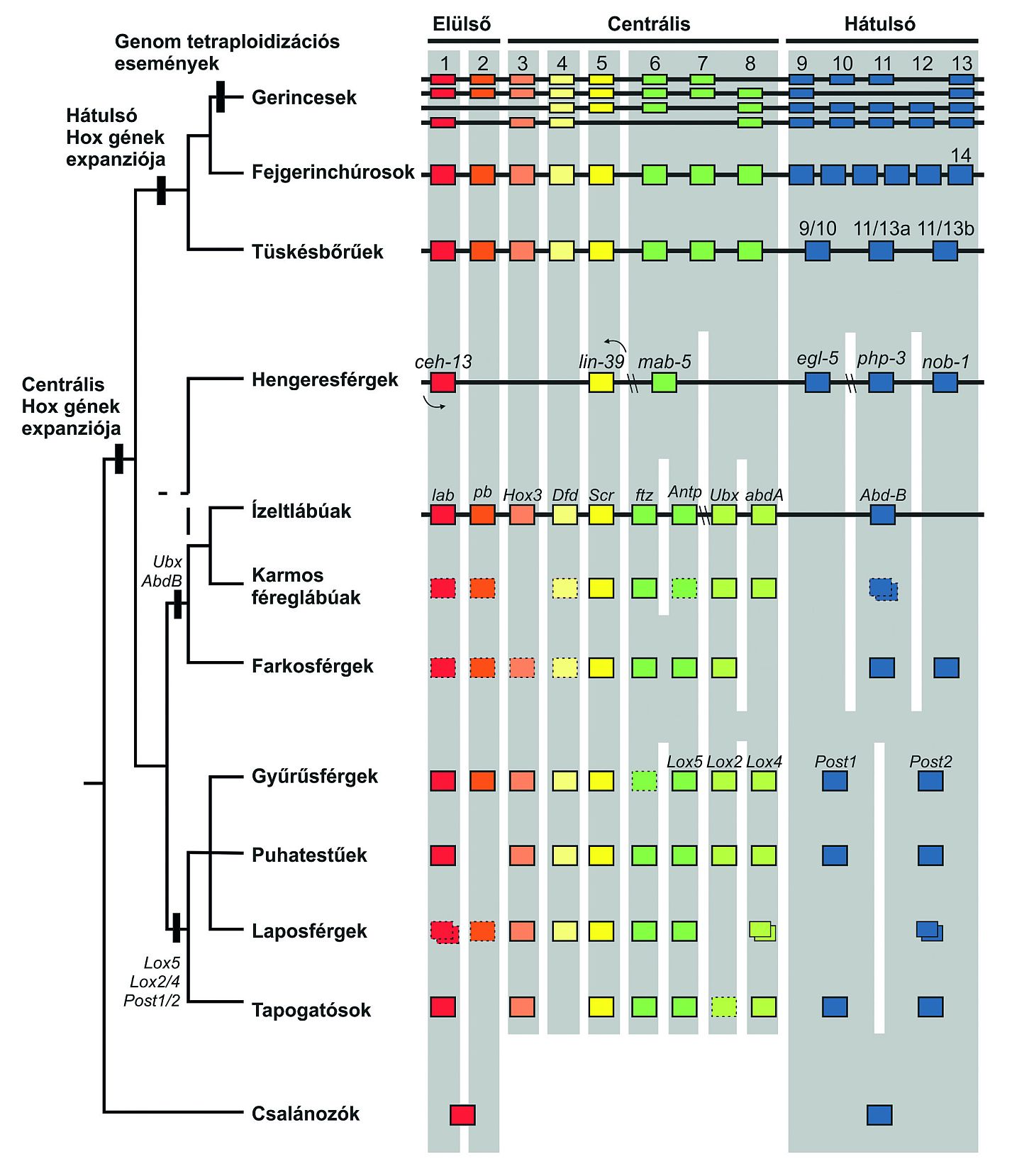
Parahox és Hox komplexek
A genomelemzések arra utalnak, hogy Metazoák/Többsejtű állatok korai időszakában egy 3 vagy 4 génből álló ősi ProtoHox komplex duplikációja két "testvér" komplexet eredményezett, a Parahox és a Hox komplexeket, ugyanis mind a Parahox komplex (fent), mind a Hox komplex (alul) jelen van, mint szorosan kapcsolt elrendezés, a lándzsahalnál (Amphioxus). A Hox komplex génjeinek száma gének duplikációjával és divergenciájával gyarapodott a Bilaterák radiációját megelőzően ill. közben.
Rovar Hox proteinek új fejlődési funkcióinak evolúciója
Rovarok evolúciója során az ősi zen és ftz Hox gének új, fejlődést szabályozó szerephez jutottak a dorzoventrális tengely képzésében (zen) és szegmentációjában (ftz), elvesztették azonban az ősi anteroposzterior tengely "regionális" mintázatát szabályozó szerepüket. A diptera ágon a zen gén duplikációja révén jött létre a bicoid gén.
Az Ubx fehérje egy új, rovarok abdominális végtagjainak repressziójához köthető motívumhoz jutott, így döntő szerepe volt a rovar testterv kimunkálásában.
Tanulságok a Hox-gének tanulmányozásából:
-
Egy kicsi, ősi Hox-klaszter korai duplikációja vezethetett a ParaHox-komplex kialakulásáshoz.
-
Tandem duplikációs események révén a paralógok száma emelkedett a Hox-komplexen belül
-
Sok Hox-gén homeodoménje divergált, másoké rögzült (valószínűleg átalakulása korlátokba ütközik, idegen szóval „constrained”)
-
A Vertebrata alapjánál genomi vagy széles skálájú duplikációk vezettek a komplexek számának növekedéséhez.
-
Több rovar Hox-génnél a homeodoménben bekövetkező gyors változások velejárói voltak az új fejlődési funkcióknak
A Hox-gének száma 2-ről 7-re nőtt a diploblaszt → bilaterálisan szimmetrikus triploblaszt átmenet idején. Ezen periódus alatt kiemelkedően fontos változások játszódtak le a testszerveződés szempontjából:
-
Kialakult a korai osztódások szabályos mintázata
-
Folyamatos bélcsatorna formálódott
-
Megjelent a mezoderma és cölóma
-
Különbözővé vált az A/P és D/V tengely
A gerinceseknél (Vertebrata) sok Hox-gén, több egyéb toolkit gén van jelen, ami jelentős redundanciát sejtet, de a duplikált gének általában megmaradtak!
-
Pl: Sonic hedgehog, Indian hedgehog, Desert hedgehog: egérben különböző szövetekben fejeződnek ki
A gerinceseknél az anatómiai és fejlődési komplexitás vitathatatlanul nőtt, ami megnyilvánul abban, hogy több sejttípusuk, jobb idegrendszerük van elődjeiknél, így nem meglepő, hogy a devontól (több mint. 300 millió évvel ezelőttől) domináns szerepük van a legtöbb földi ökoszisztémában.