A legteljesebb fosszilis anyagot a kemény részekkel, rendelkező tengeri élőlények hagyták hátra. Héjaikon és vázaikon buja paleontológiai szakirodalom sarjadt ki, aminek egyik bravúros elemzője Jack Sepkoski, aki a nyolcvanas évek első felétől a kilencvenes évekbe terjedően, hatalmas átlátó képességről téve tanúbizonyságot, 4000 vázzal rendelkező tengeri család és 20 000 nemzetség sorsát vizsgálta a Fanerozoikum 542 millió évében. A családok diverzitása a kambriumban és ordovíciumban gyorsan növekedett, a paleozoikum további részében stagnált, majd a mezozoikumi és kainozoikumi folyamatos emelkedés után a kései harmadidőszakban újabb tetőzés alakult ki.
Ezt az általános mintázatot a diverzitás csökkenései szakítják meg, amelyek tömeges kihalásoknak köszönhetők. A mintázat a különféle korrekciók után is szilárdnak tűnik, bár a régi és új idők mintázatainak eltérő jellegét sokan vitatják.
A diverzitás a szárazföldön is változott. A rovarcsaládok száma folyamatosan emelkedik a perm óta. Az edényes növények és a madarak, valamint az emlősök szerepe drámai a középső- kréta óta zajló diverzitásnövekedésben.
Általában elmondható, hogy az összes valaha is élt faj 99%-a eltűnt. Oka az, hogy kihalások mindig voltak, bár a kihalási ráták drámai módon fluktuáltak. Egyes csoportok kihalhattak, míg más csoportok szaporodtak.
A Fanerozoikum során általánosan megfigyelhető a fajok keletkezési és kihalási rátájának fluktuációja. A triász óta megfigyelhető, hogy a vízben élő állati taxonok keletkezési rátája nagyobb, mint a kihalási rátájuk: a mezozoikumban és a Kainozoikumban így a tengeri állatvilág diverzitása nőtt. Az új családok keletkezési rátája az állati evolúció során a kövületek alapján a kambriumban és ordovíciumban, valamint a nagy permi kihalást követően a korai triászban drasztikusan megnőtt.
A kihalási ráták drámaian különböztek a földtörténet során. Markánsan különbséget lehet tenni azok között a rövid időszakok között, amikor nagyszámú taxon hal ki, ilyenkor tömeges kihalásról (mass extinction) beszélünk. Ezzel szemben vannak olyan periódusok, amikor „átlagos szintű” a kihalás (background extinction).
A tömeges kihalások időről-időre meghatározó szerepet játszanak a Föld életében. Öt-hat esetben öltött a kihalás vitathatatlanul jelentős méreteket, míg több olyan periódus volt, amikor nem következett be akkora méretekben, hogy az mindenki számára egyértelműen tömegesnek számítson. Sok paleontológus véli úgy, hogy az elmúlt 600 millió évben a kihalások 26 millió évente jelentkeztek. Miután máig sincsen megegyezés az okokban egyetlen kihalás esetében sem, egyelőre egységes elmélet sem alakult ki, az mégis kirajzolódott, hogy sokkal valószínűbbek a fizikai, mint a biológiai okok.
A „klasszikus nagy” kihalások közé sorolhatóak 106. ábra:
Késő ordovíciumi - okaként a Gondwana eljegesedésének a változásait említik. Az élővilág egy stabilabb, magas tengerszinttel jellemzett periódus után sínylette volna meg a változásokat.
Késő devoni - a globális lehűlés egyes tengerek anoxiáját okozhatta.
Perm végi, vagy perm/triász (P/Tr) határon - ezt a legnagyobb, fokozatosan zajló kihalást a huzamos lehűlés okozhatta, amely a meleg, sekély tengerek regressziójával és területcsökkenésével járt.
Késő triászi - talán a csapadékmennyiség drámai növekedése okozhatta.
Kréta végi - meteorit becsapódás okozhatta.
Eocén – oligocén - fokozatos kihalás, amelyet a komoly lehűlés, eljegesedés és az óceáni áramlások megváltozása okozhatott (talán az Antarktisz körüli áramlásra vezethető vissza).
A természetes szelekció nem alakíthat ki adaptációkat olyan körülményekhez, amelyek sok évmilliós időskálán jelentkeznek, így nem várhatjuk, hogy a fajoknak a tömeges kihalások túlélésére szóló adaptációik legyenek. Hogy melyik taxon él túl, az legtöbb esetben a szerencse dolga. Nem zárható ki azonban az, hogy egyes taxonoknál véletlenszerűen kifejlődtek olyan jellegek, amelyek elősegítik jobb túlélésüket a tömeges kihalások idején.
Minden tömeges kihalás úgy változtatta meg a következő időszakot, hogy szelektíven kipusztulnak bizonyos csoportok – relatíve nő mások gyakorisága, pl. a planktonikus foraminiferák (tengeri egysejtűek) közül a tömeges kihalást csakis az egyszerűbb, nagy elterjedésű fajok élték túl. Egyébként is a kisebb geográfiai elterjedésű fajok pusztulnak ki gyakrabban az “átlagos” kihalásnál.
Másrészről a krétai kihalásnál a közeli rokon fajok csoportjai, amelyeknek összességében szélesebb az elterjedése, de az egyes fajoké szűkebb, jobban túlélték, mint azok, amely csoportoknak összességében nem olyan széles az elterjedése, de az egyes fajoké szélesebb, mint az előző csoportban. Hasonló mintázatot találtak Molluscaknál is.
Tanulság: a hosszú túlélést elősegítő jellegek sokszor különböznek azoktól, amelyek elősegítik a túlélést a tömeges kihaláskor (elterjedt csoport esetén a kis elterjedésű fajok is túlélhetnek).
A kréta végén a szárazföldön a nagytermetű gerincesek kihalási rátája sokszorosa a kistermetűeknek; a pleisztocénben is a nagy emlősök és madarak kihalási rátája nagyobb. Máskor a tengeri organizmusok haltak ki nagyobb ütemben – a szárazföldiek túlélték. Voltak olyan kihalások, amelyek mindkét környezetet érintették – a változások a két környezetben nem mindig esnek egybe.
Mind az állatoknál, mind a növényeknél is általános tendencia, hogy olyan taxonok, amelyeknél magas szintű a fajkeletkezés (speciáció), magas szintű a kihalás is, azaz magas a „turnover”. Ammoniteszeknél és trilobitáknál például az új fajok keletkezése és a kihalás is egyértelműen gyorsabban zajlik, mint például a csigáknál és kagylóknál. Az összefüggés magyarázatára több lehetséges magyarázat született:
-
Az ökológiai specializáltság foka: az ökológiai szempontból specializáltabb fajok valószínűleg sebezhetőbbek, mint a generalisták környezetük változásaival szemben. Valószínű, hogy foltszerű elterjedtségük miatt az újonnan kialakult fajok könnyebben maradnak fenn, mivel specializálódhatnak a különböző forrásokra, és elkerülhetik a versenyt más fajokkal.
-
A populáció dinamikája: a kis, vagy fluktuáló populációs mérettel rendelkező fajok különösen könnyen halnak ki. Sok kutató úgy gondolja, hogy a fajképződés esélyét is megnöveli a kis- vagy fluktuáló populációméret, bár ez a feltevés vitatott (ld. Makroevolúció)
-
Földrajzi elterjedés: A szélesebb elterjedésű fajok kisebb valószínűséggel halnak ki, mivel a helyi változások nem végzetesek rájuk nézve. Fajképzési rátájuk is alacsonyabb, valószínűleg nagyobb elterjedési/ migrációs képességük miatt.
Azok a taxonok, amelyek fajainak magas fajképzési és kihalási rátája van, „illékonyak”, diverzitásuk fluktuál, azaz drámai mértékben változik, és hajlamosak kihalni.
Foote nyomon követte a fajkeletkezés (S) és kihalás (E) változásait a Fanerozoikum 107 időszakában. Elemzései a következő eredményekre vezettek: A diverzitás jobban nőtt, ha S nőtt, és jobban csökkent, ha E nőtt. Ami sokkal érdekesebb: A paleozoikumban E hatása erősebb, mint S-é, míg S hatása jelentősebb a mezozoikumban és Kainozoikumban a diverzitás változására. Kutatásai megerősítették azt az elméletet is, hogy a diverzitás-függő faktorok, mint például a fajok közötti kompetíció stabilizálják a diverzitást egy egyensúlyi állapotban.
Sepkoski (1948-1999) a tengeri állatfosszíliák diverzitása alapján három nagy evolúciós tengeri faunát különített el: kambriumi-, paleozoikumi- és modern fauna. 106. ábra
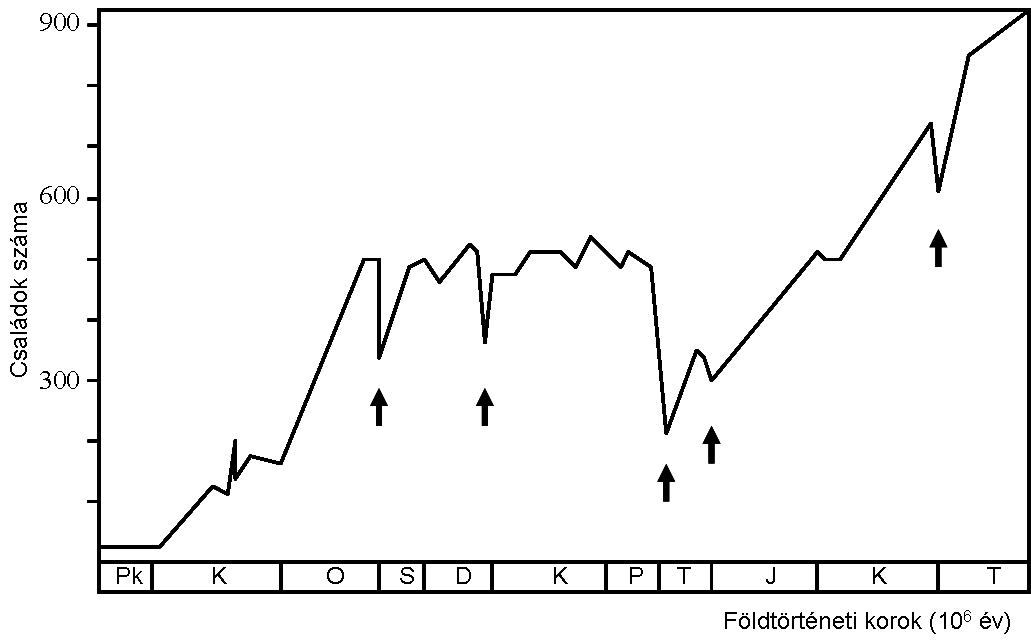
A diverzitás ugrásszerűen nőtt meg a három „nagy evolúciós fauna” megjelenésével, de az ábrán jól látszik (nyíllal jelöltük), hogy öt alkalommal a növekedési trendet mély hullámvölgyek törik meg (ordovícium, devon, perm, triász, kréta). Ezek az események a tömeges kihalások, melyek során széles földrajzi elterjedésű, magasabb rendszertani csoportok szenvedtek az átlagosnál nagyobb kihalást.
A kambriumi robbanáskor (~530 millió éve) minden ma élő nagyobb csoport megjelent (a már kihalt törzsekkel egyetemben). A paleozoikumi és triászi „explozió” során drasztikusan nőtt a családok, rendek és fajok száma, de új törzs nem alakult ki! Az új fajoknál csak a testtervek (az anatómiai képletek/szervek elrendeződése egy monofiletikus taxonon, alapértelmezésben törzsön belül) módosultak.
Honnan a különbség a két utóbbi és az első robbanás között?
Talán az az oka, hogy a kambriumi robbanás egy olyan világban történt, ahol kevés faj volt, kis méretekkel, így az ökológiai körülmények alkalmasak voltak új testtervek és új életmódok kialakulásához. Eleinte sok forma túlélt, de aztán a versengés fokozódott, új ragadozók jelentek meg és sok forma kipusztult.
Bár a Földön a másik két „élet-robbanás” idején is csekély számú faj volt jelen, azoknak sokféle testterve és életmódja miatt kis esélye volt az új törzseket létrehozó „innovációknak”.
A versengés modellezésével a faunákon belül kimutatta azt is, hogy a diverzitás változása magyarázható diverzitás-függő kompetícióval.
Az eredmények számos új kérdést vetnek fel: Miért csökkennek a fajkeletkezési és kihalási ráták a Fanerozoikum során? Hogyan emelkedhetett a diverzitás folyamatosan a perm-végi kihalás óta, ha diverzitásfüggő faktorok tendenciózusan stabilizálják a diverzitás mértékét? Milyen hatása volt a tömeges kihalásoknak az élet történetére? A továbbiakban ezekre a kérdésekre keresünk választ.
A kihalás a sorsa csaknem minden fajnak, amely valaha is megjelent, de keveset tudunk ennek tulajdonképpeni okairól. A biológusok nagy többsége egyetért abban, hogy a kihalást az okozza, hogy a faj képtelen alkalmazkodni a környezeti változásokhoz. A jelenkori populációkra vonatkozó vizsgálatok szerint messzemenően leggyakoribb kihalási ok az élőhely pusztulása, de az okok között szerepelhetnek a betelepített ragadozók, betegségek és a versenytársak is.
Ha egy faj környezete lepusztul, bizonyos populációk eltűnnek, és a faj földrajzi elterjedése csökken, hacsak korábban be nem lakott helyeket nem képesek kolonizálni. Ha a környezeti változások miatt a populációk hanyatlanak, túlélésük, sőt sokszor a faj túlélése az adaptív genetikai változáson múlik. Hogy ez elégséges-e a kihalás megakadályozására, az attól függ, hogy milyen gyorsan változik a környezet (és az optimális fenotípus) a jelleg evolúciós rátájához viszonyítva. Az evolúció sebessége függhet a rátától, amellyel a mutáció „ellátja” a genetikai variációt, és a populáció méretétől, mivel a kisebb populációkban kevesebb a mutáció. Így logikus, hogy a környezeti változás, amely csökkenti a populációméretet, csökkenti a populáció esélyét is a környezethez való alkalmazkodásra. Egy környezeti faktor megváltozása általában más faktorok megváltozását is maga után vonja, pl. a hőmérséklet változása a fajösszetételben is változást von maga után, a faj túlélése legtöbbször sok sajátság evolúciós változását kívánja meg.
Vitathatatlan, hogy mind biotikus, mind abiotikus tényezők játszhatnak szerepet a kihalásban. A csigák és kagylók elsősorban az északi tengerekben történt pliocénkori fokozott kihalásáért minden bizonnyal a hőmérséklet csökkenése a felelős.
Ésszerű feltételezni, hogy az élőlények vonalai az idő múlásával egyre jobban alkalmazkodnak, és egyre inkább ellenállnak a kihalásnak. Az evolúcióelmélet nem jósol ilyenfajta javulást (az evolúciós korlátokról szóló fejezetben részletesen ismertetjük Ernst Mayr álláspontját arról, miért nem vezet tökéletességhez az evolúció). A természetes szelekció „korlátozottsága” elvi szinten is nyilvánvaló: nincsen előrelátási képessége, így nem készítheti fel a fajt a környezet változásaira, különösen azért, mert a fajra nézve veszélyes folyamatok sokfélék, így a „kihalás rezisztencia” mint általános fogalom nehezen értelmezhető. Mindent egybevetve azt várhatjuk, hogy t időpontban egy faj (vagy akár magasabb taxon) kihalásának valószínűsége ugyanakkora, akár „régi”, akár „új” a t időponthoz viszonyítva.
A taxonok kihalása elemezhető, ha felvesszük a taxonok számát a kihalásukig eltelt idő (tkp. „koruk kihalásukkor”) függvényében. Ha a kihalás valószínűsége független a kortól, azoknak a taxonoknak a száma, amelyek nagyon magas kort érnek meg, exponenciálisan csökken. Logaritmikus ábrázolásban a görbe egyenes lesz. Ha a korral nő az ellenállás a kihalás okaival szemben, konkáv görbe adódik, hosszú „farokkal”. Van Valen hasonló elemzései (1973) egyenes lefutású görbékhez vezettek, ami arra utal, hogy a kihalás valószínűsége durván állandó, ami logikusan következik abból, hogy az élőlények folyamatosan szembesülnek olyan környezeti változásokkal, amelyek a kihalás veszélyét hordják magukban. Van Valen úgy érvelt, hogy egy taxon környezete más taxonok evolúciójának következtében pusztul le. Ő dolgozta ki a nagy hatású Vörös Királynő hipotézist (Red Queen hypothesis), amely szerint minden fajnak olyan gyorsan kell fejlődnie, ahogy csak tud, ahhoz, hogy túléljen, mert versenytársai, ragadozói, parazitái is folyamatosan fejlődnek (Az elnevezés Lewis Carroll Alice Csodaországban című művének szereplőjéről kapta a nevét, aki olyan gyorsan fut, ahogy csak bír, hogy helyben maradhasson). Az esély pedig, hogy a faj sikertelen lesz a „rohanásban”, durván mindig ugyanaz.
Kétségtelen ugyanakkor, hogy az alap kihalási ráta (background extinction) a Fanerozoikum alatt csökkent. Egyik magyarázat a jelenségre azt veszi alapul, hogy a családokon belül nőtt az átlagos fajszám. Ez a növekedés csökkenti a kihalási rátát, mivel a nagy család nehezebben hal ki, mint egy kisebb család. Egy másik lehetőség, hogy azok a magasabb taxonok, amelyek sajátságaiknál fogva hajlamosabbak a kihalásra (diszperziós képességek, élőhely) a Fanerozoikum elején kihaltak. A paleozoikumban domináns csoportok, mint a tengeri liliomok (Crinoidea), a Brachiopodák, jellemzően nagyobb kihalási rátájúak voltak, és fajaik gyorsabban cserélődtek, mint a paleozoikum utáni időket uraló kagylók, csigák, és más taxonok.
A korábban felsorolt tömeges kihalásoknak meghatározó szerepük volt a földi élet alakulásában. A perm-végi kihalás volt a legdrasztikusabb, a tengeri családok 54, nemzetségek 84, a fajok 80-90 százaléka halt ki. A szárazföldön nagy változások történtek a növénytársulásokban, eltűnt néhány rovarrend, a domináns kétéltűek, a domináns Therapsida hüllőket új Therapsidák (ide értve a mai emlősök őseit is) és diapsidák helyettesítették (köztük a dinoszauruszok őseivel). Az érintett taxonok számának tekintetében a második legnagyobb tömeges kihalás az ordovícium végén történhetett. Sokkal híresebb, de súlyosságában az előbbiektől elmarad a K/T esemény, vagy krétavégi kihalás, ekkor sok tengeri és szárazföldi növény és állat tűnt el, amelyek közül kiemelkednek a dinoszauruszok, és amelyek közül csak a madarak maradtak fenn.
A tömeges kihalások okai: A legdrasztikusabb tömeges kihalás, a perm végi kihalás okairól máig komoly viták folynak. A vitatott gyorsasággal lezajló kihalási hullámmal egybeeső rendkívül aktív vulkánosság a legnépszerűbb magyarázat (nem kizárt, hogy meteorit becsapódás válthatta ki). A vulkánosság mértékére jellemző, hogy a láva Európányi területet fedett le a mai Kelet-Oroszországnak megfelelő területből (a formációt szibériai csapdákként (Siberian traps) szokás említeni). A kitörések elég hőt termelhettek ahhoz, hogy globális felmelegedés legyen a Földön, ami megváltoztathatta az óceáni áramlásokat, talán teljes oxigénhiányt okozva a mélyebb vizekben. A globális felmelegedés hatalmas mennyiségű metán felszabadulását is okozhatta, egy pozitív felmelegedési spirált okozva Földünkön, amely, Benton és Twitchet (2003) baljóslatú szavaival: „közel volt a teljes megsemmisüléshez”.
A leghíresebb kihalási folyamat azonban kétségtelenül a K/T kihalás, kétféle okból: a Föld történetének talán leglátványosabb időszakának, a dinoszauruszok korának vetett véget, és okaira Walter Alvarez (1980; University of California, Berkeley) javasolja az elméletet, melynek alapja az, hogy az irídiumnak viszonylag magas koncentrációja jelenik meg azokban a vékony kőzetrétegekben, amelyek a kréta és harmadidőszak határára esnek. Az irídium nagy mennyiségben van jelen bizonyos meteoritokban, de nagyon ritka a Földön.
Alvarez igazán újszerű hipotézist javasolt: egy meteorit vagy aszteroid becsapódását, ami elég erőteljes volt ahhoz, hogy porral töltse meg az atmoszférát, elsötétítse az eget, lecsökkentse a hőmérsékletet, mindebből következően drasztikusan redukálja a fotoszintézist.10 km átmérőjű meteorit ütközött a Földdel 72.000 km/h-val ® nagy tüzek, szökőár, rengeteg porfelhő – körülvette a Földet – napfényt elzárta ® földi hőmérséklet lecsökkent ® a leszálló por ® irídiumréteg. A geológusok körében egyetértés van annak tekintetében, hogy ilyen esemény történt, a Yucatan-félsziget melleti Chicxulub-kráter jó jelölt a becsapódás helyére.
Sok vita: sokak szerint nem volt olyan gyors a kihalás, évmilliókig is eltartott – ellentmond a meteorit elméletnek. Talán egyes vulkánkitörések is generálnak irídiumot?
Sokat finomodtak a módszerek, és talán több érv szól a meteorteória mellett.
180 km Æ kráter a Yukatán-félsziget északi partján, ami 65 millió éve keletkezett, ma sokan úgy gondolják, hogy nem ez a becsapódás volt a döntő, hanem egy 300 000 évvel későbbi. Emellett szól, hogy 300 000 évvel a yukatáni becsapódás után még találtak dinoszaurusz kövületeket. A legtöbb paleontológus egyetért abban, hogy ez a becsapódás okozta a tömeges kihalást, bár felvetették, hogy sok taxon kihalása túlságosan elnyúlt ahhoz, hogy a korábban sejtett módon egy eseménynek legyen köszönhető, és mások úgy gondolják, hogy több környezeti változás kölcsönhatása okozhatta a kihalást. Valószínűleg gyors volt a kipusztulás
Konszenzus még nincsen, de sok minden szól a meteorit-teória mellett.
Kihalók és túlélök: A tömeges kihalások „szelektívek” abban az értelemben, hogy egyes taxonok nagyobb valószínűséggel halnak ki, mint mások. A perm-végi kihaláskor a szélesebb földrajzi és ökológiai eloszlással rendelkező, nagyobb fajszámú csiganemek nagyobb eséllyel éltek túl. A túlélés más sajátságok tekintetében véletlenszerű, például a táplálkozási mód nem befolyásolja. A szelektivitási mintázat nagyban hasonlított a tömeges és átlagos kihalással jellemezhető periódusokban: a csigáknál, és más vizsgált taxonoknál is jellemző volt, hogy a szélesebb elterjedésű taxonok kihalási rátája alacsonyabb, mint a kisebb területen élőké.
A kréta-végi kihalás túlélési mintázatai azonban drámaian eltértek a „normál” időkétől: Az alapszintű kihalások idején a csigáknál és kagylóknál a kései krétában nagyobb mérvű volt a túlélés aránya a planktonikus fejlődésű fajok között, melyeknek lárvái jól terjedtek az áramlásokkal, és a nagy fajszámú nemzetségek is jobban éltek túl, különösen, ha nagy volt a földrajzi elterjedésük. A kréta végi kihalásnál a mintázatok eltérőek: a planktonikus és nemplanktonikus lárvával rendelkező fajok kihalási rátája azonos, és a nemek kihalási rátájára nincsen hatással a fajszámuk, bár a szélesebb elterjedés előnyös. A tömeges kihalásokkor a túléléssel más jellegek korrelálhatnak, mint „átlagos” időkben.
A tömeges kihalásokkor olyan taxonok, amelyek egyébként kiváló adaptív minőséget képviseltek, kihalhattak, mert nem voltak meg bizonyos kritikus, túlélést segítő sajátságaik az ilyenkor fennálló körülmények között. Az „átlagos” időkben elkezdődő evolúciós trendek kezdeti stádiumban derékba törhetnek: szép példa, hogy bizonyos triászbeli csiga vonalakban kialakult a képesség, hogy kifúrják a kagylókat és így jussanak hozzá húsukhoz. A vonal aztán kihalt, az egyébként ígéretes jelleg elveszett, és eltelt 120 millió év, amíg, egy másik vonalban megjelentek az osztrigákat megfúró vonalak. Egy potenciálisan adaptív radiáció alapjául szolgáló jelleg így hamvába holt.
A tömeges kihalások után a fizikai és biotikus környezet nagyon valószínű, hogy komoly mértékben megváltozik. Talán ez az oka, hogy sok csoport még jóval a kihalási esemény után is zsugorodik, míg mások, sokszor korábban jelentéktelennek számító csoportok diverzifikálódnak. A diverzitás korábbi mértéke évmilliók alatt áll helyre, szélsőséges esetben, mint a perm végi kihalás után 100 millió évet vett igénybe, míg a diverzitás elérte a katasztrófát megelőző szintet.
A tömeges kihalások, különös tekintettel a perm végi és a K/T kihalások, drámai hatással voltak a földi élet további történetére, mivel újra megnyíltak az életterek. Stephen Jay Gould a nyolcvanas évek közepén azt javasolta, hogy az evolúciós változások „körei” („tiers”), tkp. konfigurációk különíthetők el, és minden ilyen megértésére szükség van az evolúció történetének teljes megértéséhez. Az első kör a populáción és fajon belüli mikroevolúciós változásoké. A második kör a „fajszelekció” szintje, a fajok differenciális proliferációját és kihalását foglalja magába átlagos kihalási ráták esetén; ez a szint befolyásolja a különböző vonalak viszonylagos diverzitását. A harmadik kör a bióta formálása tömeges kihalással, amely különböző taxonokat tüntet el, és megtisztítja az élet színpadát lehetővé téve az új evolúciós radiációkat, vadonat új irányba terelve az életet, amely csaknem független az addigi történésektől.
Bambach (2002) három funkcionális kritérium szerint osztályozta a Fanerozoikum állatnemzetségeit: Passzívak (nem végeznek aktív helyváltoztató mozgást) vagy aktívak (motilisak), fiziológiai szempontból „puffereltek”, azaz fiziológiai folyamataikat szabályozásuk mennyire tartja szűk határok között (jól fejlett kopoltyúik és keringési rendszerük van, mint pl. a rákoknak), ill. nem „puffereltek” (pl. tüskésbőrűek), és hogy ragadozók vagy nem ragadozók. A háromféle funkcionális osztályozás alapján az alternatív jellegekkel rendelkező taxonok aránya akár 200 millió éves időszakokban is stabil maradt, még akkor is, ha a globális diverzitás és a tengeri fauna összetétele erőteljesen változott. Az egyik stabil összetételből (konfigurációból) a másikba való áttolódás az ordovícium végi, perm végi, kréta végi kihalással valósul meg, ami arra utal, hogy a régóta fontos szerepet betöltő taxonok kihalása lehetővé teszi új közösségi formák létrejöttét.
Egyes csoportok eltűnése lehetővé teszi mások felvirágzását: ez a viszonylag egyszerű összefüggés írja le a tömeges kihalások hatását az új taxonok kialakulásában és diverzifikációjában.
A következőkben választ keresünk két alapvető kérdésre: (1) Miért emelkedik a diverzitás nagyobb mértékben, és eltérő időszakokban, bizonyos taxonokban, mint másokban, (2) Miért van tendencia a diverzitás emelkedésére a perm végi kihalás óta?
A diverzifikációt elősegítő főbb faktorok között van: a kompetíció alól való felszabadulás, az ökológiai divergencia, a koevolúció, és a provincialitás.
Felszabadulás a kompetíció alól
A jelen és a múlt élőlényeinek tanulmányozása közhellyé tette a megállapítást: a vonalak akkor diverzifikálódnak leggyorsabban, ha új ökológiai lehetőségekhez, „ökológiai térhez”, üres, más élőlények által el nem foglalt nichekhez jutnak. Sok lakatlan szigeten, vagy elzárt élőhelyen néhány faj kolonizálja a területet, diverzifikálódik, és olyan ökológiai nicheket tölt meg, amilyeneket máshol esetleg teljesen más rokonsági körbe tartozó fajok népesítenek be. Szép példái az ilyen adaptív radiációknak a kelet-afrikai nagy tavak cichlidái, a Galápagos-szigetek Darwin-pintyei. A taxonómiai szempontból szegényesebb szigeteken viszonylag gyakoribbak az olyan élőlények, amelyek szokatlan életformákat képviselnek. A lepkék hernyói szinte kivétel nélkül növényevők, de a Hawaii-szigeteken az Eupithecia nemzetség hernyói ragadozók. Az ennyire különleges példák általában az olyan helyeken tűnnek fel, ahol a fajdiverzitás csekély, így adaptációjuk kialakulásának kezdetibb szakaszában, mikor még kevésbé hatékonyak, nem találkoznak erőteljes versenytársakkal.
A fosszilis anyagban sok olyan példa lelhető fel, amikor egy csoport redukcióját vagy kihalását szorosan követi, vagy akár kíséri egy másik, ökológiailag hasonló csoport virágzása: a fenyők és más nyitvatermők hanyatlottak, ahogy a zárvatermők diverzifikálódtak, vagy az emlősök radiációja akkor történt, mikor (a madarak kivételével) a dinoszauruszok kipusztultak.
A hasonló jelenségek magyarázatára többféle hipotézis született, kettő közülük az érintett kládok fajai közötti versengésre épül. Egyrészről: a későbbi csoport versengéssel okozhatja a korábbi kihalását, a folyamat neve kompetitív helyettesítés (competitive displacement). A másik elmélet szerint a régi, ökológiailag beágyazódott taxon megakadályozza az ökológiai szempontból hasonló taxon diverzifikálódását, eltűnése nyomán üressé válik egy niche, lehetővé válik a másik taxon radiációja. A folyamat neve „inkumbens (hivatalban lévő) helyettesítése” („incumbent replacement”). Rosenzweig és McCord (1991) érvelése szerint a későbbi taxonnak akár kiváló adaptációi lehetnek, akkor sem biztos, hogy a korábbit helyettesíteni tudja. A két modell között nem könnyű különbséget tenni a gyakorlatban: a fosszilis anyag természeténél fogva általában nem elég finom felbontású. A fajok közötti versengés bizonyosan befolyásolta a diverzitás változásait (Sepkoski, 1996) A diverzitás növekedési rátája sokkal nagyobb volt akkor, amikor a diverzitás különösen alacsony volt, akár a kambriumban, akár a tömeges kihalásokat követő időkben. Voltak olyan időszakok (pl. a kései paleozoikum), amikor több, mint 200 millió évig a diverzitás stabil szinten volt. Hogy pontosan hogyan hatott a versengés, nem egyértelmű.
A kompetitív helyettesítés, úgy tűnik, viszonylag ritka lehetett (Benton, 1996). Ilyenkor a következő mintázat várható el:
a korábbi és a későbbi taxonok ugyanazon a helyen, ugyanabban az időben éltek
ugyanazon forrásokat használták
a korábbi taxont nem érintette jelentősen egy kihalás
a későbbi taxonba tartozó egyedek mennyisége (abundanciája) és diverzitása nő, ahogy a korábbié csökken
A hajtásos növényeknél, amelyek biztosan versengenek a térért és a fényért, pontosan ezt a mintázatot mutatták a kréta időszak folyamán, amikor a virágos növények diverzitása és abundanciája nő a virágtalanok, elsősorban a spórával rendelkező növények, mint pl. a páfrányok rovására. Hasonlóan szépen támasztja alá a kompetitív helyettesítést a kagylók és brachiopodák esete a perm végi kihalás után.
Az „inkumbens helyettesítése” valószínűleg sokkal gyakoribb folyamat, mint a kompetitív helyettesítés”. A méhlepényes emlősök nagy radiációját a Kainozoikumban általában az emlősöket addig a legtöbb tekintetben elnyomó (nem madár) dinoszauruszok és más nagytermetű hüllők kréta végi kihalásával magyarázzák.
Talán a legszebb példát az „inkumbens helyettesítésére” az ismételt helyettesítések adják. A teknősök legősibb csoprtja az Amphicelydiák???, amelyek nem tudják visszahúzni fejüket és nyakukat a páncéljuk alá. A nyakukat és fejüket behúzni képes modern teknősök két csoportja a világ különböző pontjain négy-öt alkalommal helyettesítette az Amphicelydiákat, különösen a K/T esemény után. A modern csoportok nyilvánvalóan nem tudtak elterjedni, amíg a régi jelen volt. A helyettesítés aztán párhuzamosan ment végbe a világ különböző pontjain, valószínűleg a versengés alól való felszabadulásnak köszönhető.
Ökológiai divergencia
A kulcs adaptáció (key adaptation) olyan sajátság, amely lehetővé teszi, hogy a vele rendelkező élőlény, gyakran egy új forrás vagy élőhely felhasználásával, egy új ökológiai niche-t foglaljon el. A fogalomhoz elválaszthatatlanul hozzákapcsolódik az a lehetőség, hogy az adaptáció lehetővé teszi a csoport diverzifikációját is. A csoport elfoglalhat egy adaptív zónát, hasonló ökológiai niche-k egy készletét. A rovarevő és gyümölcsevő denevérek, amelyek éjszakai életmódot folytatnak, két adaptív zónát foglalnak el, amelyek különböznek a nappali rovar-, és gyümölcsevő madarakétól. Az ökológiai tér fogalma nagy vonalakban megegyezik az adaptív zónák készletével.
Az új források és életterek felhasználási képességének evolúciója biztosan alapvetően hozzájárult a diverzitás időbeli növekedéséhez.. A tengeri sünöknél (Echinoidea) a korai mezozoikumban három rend diverzitása emelkedett nagy mértékben. Az Echinacea rendnél erősebb állkapcsok alakultak ki, így többféle táplálékot fogyaszthatnak. A szívsünök??? (Atelostomata) és a homokdollárok (Gnathostomata) ásó életmódra, a fínom szerves üledék fogyasztására specializálódtak. Az élőhely- és életmódváltást lehetővé tevő kulcs adaptáció utóbbiak esetében a lapos testforma, és a különlegesen módosult csőlábak (???), amelyek képesek a szerves üledék begyűjtésére és a szájba juttatására. Bambach (1985) szerint a tengeri állatok egész Fanerozoikumban zajló diverzitásnövekedése magyarázható az ökológiai tér a homokdollároknál tapasztalhatóhoz hasonló evolúciós újítások segítségével való betöltésével. Az új életterekbe való behatolás, és új táplálkozási szokások felvétele magyarázza a négylábú gerincesek legtöbb családjának diverzifikálódását is, mint az a békák, kígyók, madarak esetén szépen nyomonkövethető (Benton, 1996).
A magyarázat a homokdollárok diverzifikálódására legalábbis plauzíbilis módon a kulcsadaptációik számlájára írható, de a szigorú természettudományos tesztet nem állná ki. Jóval erősebb bizonyítékok nyerhetők olyan esetben, ha a diverzifikáció egy adott jelleggel korrelál, és ez a jelleg függetlenül kifejlődött több különböző kládban. Ilyen tesztek ma is élő csoportok esetén végezhetők. Egyes, az új jelleggel rendelkező kládok diverzitása összevethető testvér??? csoportjaikkal, amelyeknél a jelleg ősibb állapota van jelen. Miután a taxonok egyidősek, fajszámaikban tapasztalható különbség a magasabb diverzifikálódási rátának köszönhető, nem a kornak. Ha a párhuzamosan kifejlődő jelleg konzisztensen magas diverzitással jár, hipotézisünk, miszerint a jelleg okozza a magasabb diverzifikációs rátát, támogatást nyer.
Több csoport alkalmazta az „ismételt testvér-csoport összehasonlítás” („replicated sister group comparison”) módszert növényevő rovarokra és növényekre. A zöld növények vegetatív szöveteivel való táplálkozás legalább ötvenszer kifejlődött a rovaroknál, elsődlegesen ragadozó és detrituszevő ősökből. A filogenetikai analízis 13 növényevő klád nem-növényevő testvércsoportját azonosította. A növényevő testvércsoportnak több faja volt. Ez a jelentős korreláció támogatja a hipotézist, hogy a növényevő adaptív zónába való belépés segítette a magasabb diverzitás kialakulását. A további vizsgálatok arra irányultak, hogy olyan növényi kládokat vizsgáltak, amelyek gyantával, vagy gumiszerű anyagokkal védik magukat a növényevő rovarok ellen, és rendelkeztek ilyen anyagokat nélkülöző testvércsoporttal. A védőanyagokkal rendelkező taxonok 16-ból 13 esetben több fajjal rendelkeztek, így lavonható a következtetés, hogy a védelmet szolgáló anyagok elősegítették a diverzitás növekedését a vizsgált csoportokban.
Modern csoportok tanulmányozása során tudottá vált, hogy a diverzitás nagyrészt olyan, közeli rokon fajoknak köszönhető, amelyek úgy kerülik el a versengést, hogy egészen finom különbség van közöttük a források használatában. A paleotológusok is arra a következtetésre jutottak, hogy a nichek felosztása egyre finomabb skálán zajlott: az egyes, általában helyi közösségeket tükröző lerakódások több fajt tartalmaznak az újabb, mint a régebbi rétegekben. A fajszám növekedése nagyobb, mint a főbb formák, vagy adaptív zónák növekedése, ami arra utal, hogy az egyre növekedő számú faj úgy maradhat fenn együttesen, ha a hasonló forrásokat egyre finomabban osztja fel egymás között.
Koevolúció
A fajok közötti kölcsönhatások többféle módon támogatják a diverzitás evolúcióját. A fajok forrásként szolgálnak más fajoknak, így egyik csoport diverzebbé válása támogathatja egy másik csoport diverzifikációját. A 700 fügefaj a különböző megporzó fügedarazsak egyetlen táplálékforrása, a darazsaknak pedig megvan a maguk fajspecifikus hengeresféreg-parazitájuk.
A ragadozók és zsákmányállatok koevolúciója szintén növelheti a diverzitást. A mezozoikum tengeri forradalmában mind a ragadozók, mind a zsákmányállatok morfológiai változatossága és taxonómiai diverzitása jelentősen megnőtt. Halak és rákok számos módon alkalmazkodtak ahhoz, hogy kagylók héját bontsák fel, a kagylók pedig sokféle védelmet fejlesztettek ki az új ragadozók ellen, többek között töviseket és vastagabb héjakat.
Provincialitás
A provincialitás annak a mértéke, mennyire oszlik meg a bióta??? A különböző földrajzi régiók között. A fauna vagy flórabirodalom olyan régió, amelyben sok a megkülönböztető, helyi taxon. Korunk faunája és flórája több provinciára oszlik, mint valaha az élet története folyamán. A taxonok kozmopolita eloszlását a mezozoikum nagy részében, és a Kainozoikumban trendszerűen felváltotta a lokalizáltabb eloszlás, ami a fő oka lehet a diverzitás globális emelkedésének ugyanezekben az időszakokban.
A tengeri állatoknál a faunaprovinciák száma az egész palozoikumban alacsony lehetett, és a triász elején elérte a mindenkori minimumot, amikor a perm végi nagy kihalást túlélő taxonok annyira elterjedtté váltak, hogy egyetlen, világszerte elterjedt provinciáról lehet beszélni.
A jura, kréta, de különösen a harmadidőszak során a tengeri állatok egyre tagoltabb eloszlást mutattak: az atlanti és a pacifikus régió is egyre több, a szélesség szerint szerveződő régióra tagolódott.
A szárazföldeken hasonló tendenciák voltak megfigyelhetők: a szárazföldi gerinceseknek különböző faunái fejlődtek ki a nagyobb szárazföldeken a mezozoikum későbbi részében, és a Kainozoikumban, és a dinoszauruszok és más csoportok viszonylag széles eloszlási doménjeit követték a mai gerincesek sokkal finomabb szélességi eloszlásai.
A fenti trend alapvető oka a földtömegek eloszlásának lemeztektonikai mozgásokból eredő megváltozása. A Pangea triászban történő feltöredezése után a földtömegek pólustól-pólusig terjedése addig soha nem tapasztalt módon árnyalt klímaviszonyokat teremtett. A kontinenseknek ez az újabb elrendeződése két, egyre függetlenebb óceánrendszert teremtett, az indiai-pacifikus és az atlanti rendszert, és az óceáni keringések soha nem tapasztalt erősségű sűrűség grádienst teremtett a sarkok felé. Nemcsak egyre többféle környezet jött létre, hanem a földtömegek is fragmentálódtak, ami szintén kedvezett a divergens evolúciónak és megakadályozta a fajok közötti kicserélődést, ami, versengéssel vagy predációval, csökkentheti a diverzitást.
A környezeti változás szerepe
A klíma és más környezeti faktorok sokféle, bonyolult módon befolyásolták a keletkezési és kihalási rátákat. A klíma nagyobb mérvű változásai a kihalási ráta emelkedését, és az élőhelyek és vegetáció típusok eloszlásának megváltozását hozzák magukkal, ami a taxonok eloszlására kihatva diverzifikálódáshoz vezethet. Az új adaptációkkal rendelkező, új taxonok kialakulása legtöbbször sokkal inkább a biotikus változásokra vezethető vissza, mint a szűk értelemben vett klimatikus változásokra. A középső-eocénben például (50-40 millió évvel ezelőtt) az éghajlat hidegebb és szárazabb lett, a szubtropikus erdőségeket többnyire szavannák váltották fel, a főemlősök és más fán lakó élőlények diverzitása lecsökkent, híg a nagy növényevőké megnőtt. Mindent egybevetve azonban a harmadidőszakban a hőmérséklet változás nem korrelált szorosan az emlősök fajkeletkezési és kihalási rátájával. A kulcsinnovációk és biotikus kölcsönhatások megkérdőjelezhetetlen fontossága mellett a környezeti faktorok szerepének fontossága kérdéses.