A devont szokás a halak koraként számon tartani, de legalább ennyire jogos a „növények kora” elnevezés is. Nevét az angliai Devonról kapta. Éghajlata meleg, a tengerszint emelkedik, a kontinentális selfeken sekélytengerek jellemzőek 52. ábra
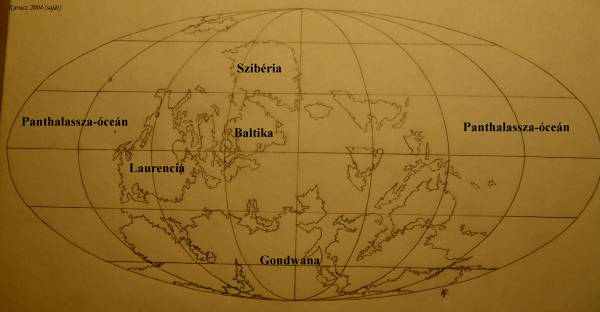
A kaledóniai hegységképződés miatt a klíma differenciáltabbá válik a Földön. A korszak jellegzetes üledéke az „Old red sandtone” vagy régi vörös homokkő. Ez a típusú üledék előfordult már a karbon elején és a szilur végén is. A vörös homokkő színét az oxidált szárazföldi üledékek adják, képződése meleg, száraz időszakokra jellemző. A devonban gyakori volt még az evaporit (szárazföldi sópárlatok) képződése is, mely a forró, sivatagos területekre jellemző. A meleg tengerekben elterjedtek voltak a sztromatolitok, a zátonyok.
"Devoni robbanás": ugrásszerűen megnőtt a jelentős fosszíliák száma, új, modern élővilág alakult ki, a szárazföld „kizöldült”. Az algák, gombák, baktériumok és az eleinte csak pár centis növények hatására kialakult az első talaj. A növényeknél megjelentek a megaspórák. Devoniak az első levél-, gyökér- és évgyűrű fosszíliák. Az ősharasztok felvirágzása és hanyatlása a devonban volt, egyesek elérték a fa termetet, megjelentek az első erdők. A meleg párás éghajlat és a növényi biomassza tömege lehetővé tette az első kőszénrétegek kialakulását. 400 millió éve a nyitvatermőkkel (Gymnospermatohyta) jelentek meg az első magvas növények is, a magvaspáfrányok.
A késő-szilur-korai-devon meleg vizeit kistermetű növények, mohák, ősharasztok szegélyezték. A középső-devonban aztán egyes növények eltávolodtak a vizektől. A tájkép szempontjából nagyon fontos változás volt a középső-devonban a fatermetű növények kialakulása (kazahsztáni leletek). A növények elágazódásai egyre bonyolultabbak lettek, tüskék jelentek meg, amelyek ellaposodhattak, levélkék alakulhattak ki belőlük. Az új mintázatok alkalmasabbak a fény hatékonyabb összegyűjtése. A hajtások egy síkban osztódhattak, majd síkjukban kialakuló szövetek kialakíthatták a nagy leveleket. A fényért nemcsak magukban az egyedek küzdöttek. A nagy magasság előnyt biztosított, ehhez viszont még erősebb test szükségeltetik. A másodlagos fás szövet (xylem) és erőteljes gyökérzet akár 50 m magasságú fák kialakulását is lehetővé tette.
A mai növényi testtervek közül valamennyi megjelenik, és 32 olyan típus is, aminek már nyoma sincsen (a zárvatermőket nem tartják külön testtervnek).
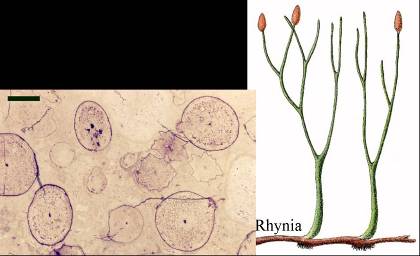
A Rhyniák 53. ábra jelentősége felmérhetetlen a növényvilág szempontjából. Végálló eusporangiumokkal rendelkező águkból alakultak ki Trimerophyták 54. ábra, a későbbi „nagylevelű” formák (Filicophyta, Cladoxylophyta), majd a „klasszikus” Macrophyllek („nagylevelűek”) ősei: a páfrányoké, „magvaspáfrányoké”, nyitva- és zárvatermőké. Erre az ágra jellemző az unipoláris, bilaterális embrió (zárvatermőknél: bipoláris), a valódi sporangiumok, és hogy a gyökér és a levél nem homológ szervek.

A laterális sporangiumokkal rendelkező ágból származó Zosterophyllophyta 55. ábra, utódai a „kislevelű” formák. Közülük a Lycophyta (korpafüvek) élnek ma is. 56. ábra


A korpafüvek egyedfejlődési mintázatának szélsőséges megváltozása (extrém peramorfózis; a sporofiton kései mintázata előretolódott az egyedfejlődés folyamán) vezetett el a zsurlókhoz (Sphenophyta). Erre az ágra jellemző: microphyllum („apró levél”, bipoláris, radiális embrió, valódi sporangiumok, a gyökér és a levél homológ szervek, azaz a gyökér módosult levél (tiszta formában a pikkelyfáknál vezették le).
A magvak előnyeit nem kell külön hangsúlyozni: védik, táplálják az utódot, ráadásul csak akkor „indítják el” fejlődését, amikor a körülmények megfelelőek. A Moresnetia ős-magvaspáfrány makrosporangiumai erősen magház jellegűek. Az Archaeosperma arnoldi az egyik első magvas növény. A Progymnospermatophyták (lásd még karbon) külön törzset alkotnak, csak anatómiájuk emlékeztet a fenyőkre, egyébként heterospórás harasztok. Benépesítették a víztől távoli, addig elérhetetlen élőhelyeket. 57. ábra
14.57. ábra - Progymnospermatophyta: Cecropsis/felső-karbon (balra), és Eospermatopteris valamint Aneurophyton germanicum sporangiumai /felső-devon/ (jobbra)
 |
Az első ősharasztok édesvizek partjain éltek, később benyomultak a szárazföldek belsejébe. Virágzásuk az alsó devonra esett, a középső devonban már hanyatlani kezdtek és a felső devon kihaltak. A felső devonban hatalmas termetű zsurlófák is éltek, köztük a Pseudobornia ursina, melynek teljes magassága a 20 m-t is elérhette. Hasonló magasságot értek el a korpafűfák is, melyek közül a Cyclostigma kiltorkense 25 m magas is lehetett. A Wattiezia a legkorábbi (385) ismert ősharaszt fa, 20 millió évvel későbbről az Archaeopteris (akár 50m).
Az első ismert tengeri gombafonal 870 millió éves. A homályos eredetű szárazföldi gombák már a szilurban jelen voltak. Jelenlétükkel segítették a talaj kialakulását és élelmet jelentettek a kisebb élőlényeknek. A talaj, a növényi maradványok, gombáknak új élőhelyet biztosítottak, akárcsak a fák, amelyek az ég felé is kiterjesztették az életteret. A devonban megjelennek a fatermetű korpafüvekkel obligát szimbiózisban élő Basidio- és Ascomycoták. Hogy ez a szimbiózis hozta ki őket a szárazföldre, erősen valószínűsíthető, mivel a ma élő összes Basidiomycota egyben fakultatív szimbionta is!!!
A Foraminiferák felvirágzása a Devonban történt. Jellemzőek a fenéklemezes korallok, tüskésbőrűek, az ammoniteszek tovább fejlődnek, a trilobiták viszont tovább hanyatlanak.

1.: Archaeosigillaria (Lepidodendropsida-Lycophyta)/középső-felső-devon/Méret: 5-10 m
2.: Heleniella (Lepidendropsida - Lycophyta) / középső-devon-alsó-karbon / Méret: 5 - 15 m
3.: Lepidosigillaria (Lepidodendropsida-Lycophyta)/középső-devon-alsó-karbon/Méret: 5 – 20 m
4.: Barrandeina dusliana (Barrandeinophytales aff. Lycophyta s. l.)/középső-felső-devon/Méret: 2 – 5 m
5.: Duisbergia mirabilis (Duisbergiophytales i.c. / aff. Lycophyta s. l.) / középső-felső devon / Méret: 3 – 8 m
6.: Pseudobornia ursina (Pseudoborniopsida - Equisetopsida) / Felső-devon / Méret: 25 m
7.: Lepidosigillaria whiter (Lepidodendropsida-Lycophyta) / középső-devon-alsó-karbon / Méret: 15 – 20 m
8.: Metaclepsidropsis (Coenopteriodopsida-Filicophyta)/felső-devon-alsó-karbon / Méret: 3 – 8 m
9.: Cyclostigma kiltorkense (Lepidodendropsida-Lycophyta) / felső-devon-alsó-karbon / Méret: 25 – 30 m
10.: Archaeopteri (macilenta) (Progymnospermatophyta) / középső-felső-devon / Méret: max. 10 m(?)
11:. Seicidium (a.l. Chara) (Charaphyta) / devon /
12.: Chovanella (a.l. Chara) (Charaphyta) / devon /
13.: Eochara (a.l. chara) (Charaphyta) / devon /
A: Parapolydesmia (nodosa a.l.?) (Archipolypoda) / felső-devon-alsó-karbon / Méret: 1,5 – 2 m
B: Ichthyostega (Amphibia) / felső-devon / Méret: cca. 1 m
C: Acanthostega (Amphibia) / felső-devon / Méret: 1 - 1,5 m
D: Panderichthys (Crossopterygii) / felső-devon / Méret: 40 - 50 cm
E: Eusthenopteron (Crossopterygii) / középső-felső-devon / Méret: 40 cm
F: Climatius macnicollii (Acanthodii) / felső-devon / Méret: 6 - 8 cm
G: Eurypterida
H: Bothriolepis (Placodermi) / felső-devon / Méret: 30 - 35 cm
I: Ventastega curonica (Amphibia s.l.) / felső-devon /
A devonban jelentek meg a pókok, atkák, ikerszelvényesek, százlábúak, és a rovarok első, röpképtelen képviselői (396: Rhynie kovakő, Skócia).Tengeri liliomok akméja (virágkora).
Az tüskésbőrűek közül teljes osztályok (Cystoidea, Carpoidea) a devon végén kihalnak.
A halak evolúciójának fontos szakaszai játszódtak le a devonban. Az időszak halairól egy skóciai lelőhely az Orcadiai-medencének elnevezett, egykor hatalmas tó által borított 5-10 cm vastag üledékekkel fedett vidékről származó kövületek árulnak el legtöbbet. A "régi vörös homokkő" (old red sandston) alsóbb rétegei rejtenek állkapocs nélküli páncélos halakat. Az tengeri skorpiók (Eurypterida) többsége a devon végi kihalás áldozata lett, karboni képviselők már csak az édesvizekben éltek. Az állkapocs nélküli halak a folyókban, tavakban fordultak elő, páncéllal rendelkeztek - védelem az édesvízi „tengeri skorpiókkal” szemben. A korai állkapocs nélküliek lassú mozgású, aljzaton élő formák, a mai ingolák ősei lehettek. Páncélzatuk később elcsökevényesedett.
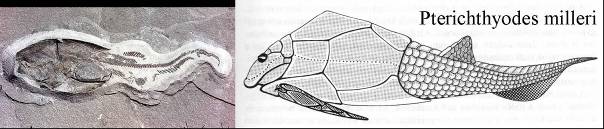
Az ordovíciumban megjelent páncélos őshalak (Placodermi) virágkora (akmé) a devonban volt. Egyik típusuk erősen páncélozott, lassú fenéklakó forma volt, másik típusuk gyors, cápaszerű. Tavakban és tengerekben is laktak. A Placodermik közé tartoztak a Pterichthyodes („szárnyas hal”) nem tagjai, köztük az emblematikus P. milleri. 59. ábra A feji része erősen páncélozott volt, míg a farki végén csak lágy részek voltak. Az elsők között írták le a 19. század elején a régi vörös homokkőből.
A Duncleosteus 10 m hosszúságú ragadozó páncélos őshal volt, hatalmas állkapcsával a korabeli tengerek legnagyobb ragadozói lehettek. Az állkapocs a kopoltyúíveket támasztó vékony csontok módosulásával alakulhatott ki. A placodermik a devonban kihaltak.
A legtöbb mai halfaj a szilur végén kialakult csontos halakhoz tartozó sugarasúszójú halak (Osteichthyes: Actinopterygii) közül kerül ki, amely a gerincesek legfajgazdagabb osztálya. Belső vázuk fokozatosan elcsontosodik, kialakul a kopoltyúfedő. Fejlett felső vállöv-csontjuk van, ganoid, cikloid, vagy ktenoid pikkelyeik lehetnek. A vörös homokkő középső rétegeiből előkerült Cheirolepis (25 cm). Úszói nagyon hasonlítottak a mai csontos halakéira. Fogait is ugyanazok a csontok hordozzák, mint a későbbi gerincesekéit. Súlyos, csontos fejének elemei igen jól mozgathatóak. Az Actinopterygidák (sugarasúszójúak) első képviselői között találjuk.
A csontos halak másik osztálya a húsos- vagy izmosúszójú halak (Sarcopterygii). Két nagy csoportjuk a bojtosúszójú halak (Coelacanthimorpha) és a Rhipidistia. (Mivel a Crossopterygii nevet szinonimájaként, alcsoportjaként is említik a Sarcopterygiinek az irodalomban, használatát jelen felsorolásban kerüljük.)
A bojtosúszújúak a kora devonban jelentek meg, virágkorukat a késő devonban és a karbonban élték. Feltételezések szerint a devontól a permig édesvízi ragadozó életmódot folytattak, melyet a triászban mélytengeri ragadozó életmódra cseréltek. Testfelépítésük nem változott jelentősen az idők folyamán: zömök test, két részből álló porcos koponya, belső orrnyílás nincs, vékony pikkelyek, bojtszerű páros úszók, trificerk farokúszó, a belső vázukat alkotó csontok homológok a szárazföldi gerincesek vázelemeivel, a fogak nem nőnek össze csontlemezzé; gerinchúr; elcsontosodó gerincoszlop. 70 millió éve (kréta) kihaltnak vélték a csoportot, mígnem 1938 decemberében Dél-Afrika keleti partjainál a Chalumna torkolatánál Courtenay-Latimer megtalálja a Latimeria chalumnae-t. A fajt az East London Múzeum munkatársa J. L. B. Smith ichtyológus írja le. A L chalumnae 1,5 m hosszú, 80 kg, szürkéskék, fehér foltokkal, bőre erősen nyálkás, agytömege 3 g. Szárazon 1 órán belül elpusztul, rendkívül érzékeny a hőmérséklet változásaira. Végtagjait minden irányban képes mozgatni. 150-400 m-es mélységben él, elsüllyedt korallok között ragadozó, halakra, rákokra vadászik. Elevenszülő, megtermékenyített petéi teniszlabda nagyságúak. Ritkaságuk alacsony szaporodási rátájukkal függ össze. A bennszülöttek ismerték, bőrét bicikligumi javításakor használták. 1997-be találták meg indonézia vizeiben a L. menadoensis-t.
A Sarcopterygii másik csoportja a Rhipidistia a bojtosúszójúakkal közel egy időben tűnt fel, de velük ellentétben elhagyták az óceánt és édesvíziekké váltak. A Rhipidistia lebenyes úszójú hal volt. Jellemzői: Három orrnyílás: 1. testfelszín – orrüreg, 2. orrüreg - szemüreg (könnycsatorna), 3. orrüreg - szájüreg (choana). Megerősödött páros úszók, dificerk farokúszó, a fogak gyűrt dentinje a kétéltűekéhez hasonló, tüdő-légzés, édesvízi élőhelyek. A Rhipidistia két csoportja a tüdőshalak (Dipnoi) és a négylábúak (Tetrapodomorpha) a devon elején különült el.
A Dipnoi virágkora a devontól a triász közepéig tartott, a triász végén többségük kihalt, csak néhány fajuk él ma is Ausztráliában, Afrikában, Dél-Amerikában. Ezek lehetővé teszik a fosszilis formák életmódjának tanulmányozását. Nem ősei a fejlett gerinceseknek (túlspecializált fogak, a végtagok eltérő belső váza). A régi vörös homokkő középső devon rétegeiből előkerült a tüdős halak közé tartozó Dipterus is. A tüdőshalak a devonban érték el evolúciójuk csúcsát.
Az első Tetrapodomorphák masszív uszonyai "preadaptívan" többszörösen kapcsolódtak a csontvázhoz.
Vermey elmélete kimondja, hogy azok a rendszertani egységek, amelyekben hatékonyabb a fajképződés, több független elemből állnak mint a fajképződés terén kevésbé hatékony taxonok. A független elemek nagy száma emeli a lehetséges megoldások számát egy adódó problémára. A Sarcopterygii csontváz is rengeteg részből áll, így nem csoda, ha a csoport nagy evolúciós ugrásokra volt képes.
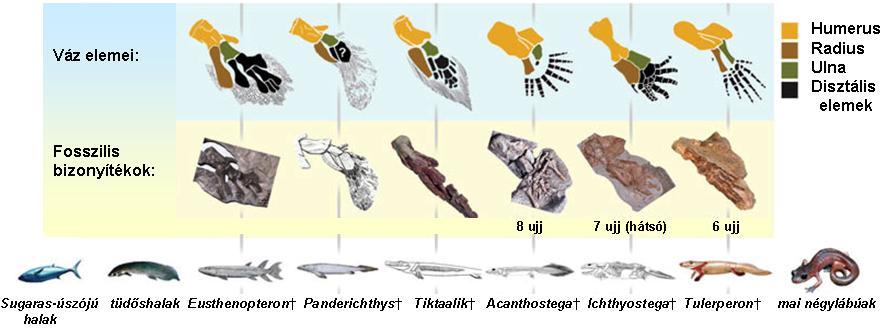
A Rhipidistia (Sarcopterygii) csoportból alakultak ki édesvizekben a mai szárazföldi gerincesek ősei a Tetrapodák kb. 400 millió éve. A vízhez alkalmazkodott élőlények álltak át szárazföldi életre. Szemben a halak oldalirányú elhajláshoz alkalmazkodott gerincével a kétéltűek gerincére elsősorban lefelé nehezedik a terhelés ® a gerinc és a hozzá kapcsolódó izomzat módosulása révén mozgáskor nem "lóg be" Þ fej tartásához új izmok kellenek Þ hasfal megerősödik a belső szervek nyomásának ellensúlyozására. Kétéltűek, hüllők oldalt hullámzó mozgása "hal-örökség" - szaggatott lépések visznek előre. A bojtosúszósok uszonyainak felépítése klasszikus értelemben vett járásra alkalmatlan, ízületeik kötöttebbek, vagy egyáltalán nem hajlanak. Az Eusthenopteron hal (~390) a bojtosúszójúak egy kihalt ága, melyeket korábban a négylábúak ősei köré soroltak hiányzó láncszemként. Az első ismert kétéltűek végtagjai, ízületei igencsak fejlettek. A vállöv levált a koponyáról, és nagymértékű erősödése mellett a csontváz egyéb részeihez szervesen kapcsolódik. A medenceöv sem csak "lóg" az izmokon. A szárazföldi négylábú állatok és az Eusthenopteron-szerű halak 60. ábra között pár évtizede a fosszíliák alapján még óriási volt a szakadék, mára viszont számos köztes fosszíliát megtaláltak, nincsenek „hiányzó láncszemek”.
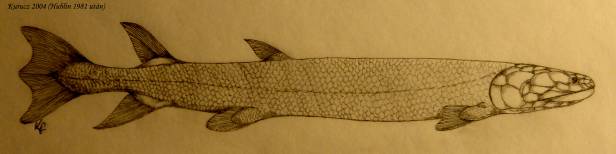
A Panderichthys (397) halnál már megfigyelhető a tetrapoda-szerű fej. A 390 millió éve élt Livoniánának csak egy állkapocsdarabját ismerjük. A Tictaalic izmosúszójú hal 375 millió éve élt. A hal és kétéltűek közötti „hiányzó láncszem” után való céltudatos kutatás során, devonkori kőzetek vizsgálatával találták meg 20 példányát Kanadában a Grönlandhoz közeli Ellesmere szigetén 2004-ben. Mivel felépítésében halakra és négylábúakra emlékeztető jegyek egyaránt felfedezhetők, ezért egyik felfedezője, Neil Shubin „fishapod” („hal-lábú”) névvel jellemzi. A Tiktaalik ízig-vérig átmeneti faj volt, keveredtek benne a halakra (pl. még jellegzetes pikkelyei voltak) és a négylábúakra jellemző tulajdonságok 61. ábra. A legfontosabb átmeneti tulajdonságot az úszók mutatják: ujjak még nem voltak megtalálhatók, de támasztékul szolgálhattak. A csuklószerű végtagok, az úszó felépítése és a feltételezett nagy izomköteg viszonylag gyors mozgást biztosított a számára a szárazföldön, DE semmiképp sem tekinthető még szárazföldi állatnak.

Ugyan még volt kopoltyúja, de azt már nem fedte csontos lemez, mint a ma is élő halfajok esetén, így képesek voltak fejüket oldalirányban is mozgatni. Ezt az először náluk megjelenő nyak is segítette. A nyakat a vállöv különítette el a koponyától. A sekély vízhez való alkalmazkodás során fejlődésnek indult a szegycsont, ami elsegíthette a szárazföldi élethez való alkalmazkodást és magának a szárazföldi életformának a kialakulását.
A 365 millió éve élt Acanthostegának 8 ujja volt végtagonként. Feltehetően édesvízben élt, mint ősei. 62. ábra

Az első valóban kétéltűnek tulajdonítható 7 ujjú Ichtyostega fosszília 375 millió éves. A kígyózó mozgású Ichthyostegának 63. ábra oldalirányban lapított teste, hajlékony gerince volt. Végtagjai a halakhoz képest eltérően kapcsolódtak, az első végtag függetlenedett a fejtől, testüket fel tudták emelni. Kopoltyúfedői voltak, feltehetően főként vízben éltek.
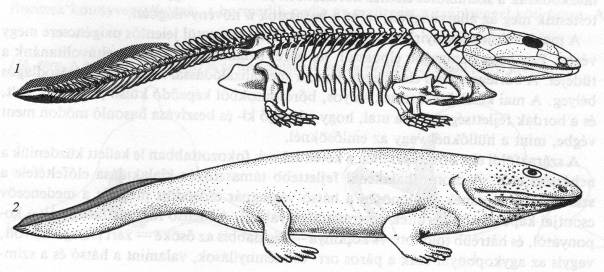
Az első szárazföldi kétéltűeknek (~360) fajtól függően 5-9 volt az ujjszáma. A 363 millió éve élt Tulerpetonnak 6 ujja volt. 64. ábra
A devonban jelentek meg a pókok, atkák, ikerszelvényesek, és a rovarok első, röpképtelen képviselői (396: Rhynie kovakő, Skócia). Az intenzív fotoszintézis miatt nőtt a légköri CO2 megkötése, amely az üvegházhatást csökkenve erős lehűlést okozott. Feltehetően ez okozta a földtörténet öt nagy kihalási korszaka közül a devonit, mely során a családok 20%-a, a genuszok 70%-a, a fajok 80%-a eltűnt. A nagy vesztesek a sztromatolitépítők, a rugosa és tabulata korallok, pörgekarúak, trilobiták, ammoniteszek, az állkapocs nélküli- és húsos úszójú halak.