Tartalom
A tradicionális neo-darwini teória megmagyarázza, hogy a természetes szelekció, genetikai sodródás, és génáramlás együttesen hatva a genetikai variáció nyersanyagára hogyan hozza létre a földi élet káprázatos sokszínűségét. De miért NEM fejlődtek az organizmusok bizonyos irányba? Miért NEM fejlődtek ki bizonyos jellegeik?
Ernst Mayr a következőkben látja annak okait, hogy az evolúció nem alkot tökéleteset:
Genotípus potenciálja: meglevő genetikai adottságok korlátozzák a potenciálisan kialakítható tulajdonságok körét. Méretkorlátok pl: törpecickánynál kisebb emlős nincs, repülő madarak tömege nem léphet át egy határt stb.
Genetikai variáció hiánya: csak bizonyos fokú variációt képes elviselni egy faj, így drasztikus változás esetén lehet, hogy pont az nincs meg, ami kell.
Sztochasztika: kataklizmák során olyan származásvonalak is kihalhatnak, melyek optimálisan adaptálódtak. A drift véletlenszerűen fixálhat adott környezetben kedvezőbb vagy kedvezőtlenebb variánsokat.
Törzsfejlődési előzmények korlátai: a struktúra megszabja a reakciót. Pl. szilárd váz előnyös: ízeltlábúak külső, gerincesek belső vázat alkottak. Ezután már az ízeltlábú vonalon jóval korlátozottabb pl. a testméret (a vedlés miatt). Vagy szárazföldi gerincesek légzőtraktusa és tápcsatornája kereszteződik, ami előnytelen, ám a bojtosúszósoktól kezdve megvan. A nervus vagus egyik ága azért ír le óriási kanyarulatot, mert eredetileg a 6. kopoltyúívet kerülte, amelynek leszármazottja az aorta pulmonaris.
Nem genetikai módosulás képessége: minél nagyobb a reakciónorma (azaz sokféle fenotípust tud létrehozni az adott genotípus a különböző környezetekben), annál jobb az alkalmazkodóképesség. Alacsonyrendűeknél általában nagyobb, de embernél is jelentős: pl. alföldi vagy magashegységi környezetben sokféle fiziológiai jellemző különbözhet, akár egyazon egyednél is (lásd: magashegyi edzőtábor). Baldwin-effektus: új, speciális környezetbe kerülve a szelekció kedvez azon géneknek, melyek megerősítik, majd helyettesítik a nem genetikai módosulás képességét. Az effektus máig vitatott.
Reagálás nehézségei a posztreproduktív szakaszban: szaporodást követően már nem (vagy alig) érinti a szelekció a betegségeket, egyéb negatívumokat (pl. időskori betegségek: Parkinson- és Alzheimer-kór, vagy a reproduktív életszakasz végén jellemző betegségek, mint az emlő- vagy prosztatarák).
Ontogenetikus interakciók: kompetitív jellegűek: az egyed szervei és struktúrái között verseny zajlik. Ha valaminek fejlődnie kell, az gyakran a másik rovására történik. Mindig visszamarad valami a feleslegessé váló struktúrákból: csökevények, maradványok (lásd pl. a troglobiontákat vagy bálnák és kígyók hátsó végtagjait).
A genotípus struktúrája: transzpozonok, intronok, repetitív szekvenciák, nem-kódoló DNS funkciója körül sok a bizonytalanság.
Vannak ÁLTALÁNOS KORLÁTOK, amelyek minden élőlényre hatnak (pl. a gravitáció hatása a morfogenezis alatt végig érvényesül), és FILETIKUS KORLÁTOK, amelyek rokon organizmusok szűkebb körére vonatkoznak. A molekuláris, esetleg mechanikai kölcsönhatások, vagy térbeli viszonyok bizonyos irányokba „terelhetnek” egyes fejlődési folyamatokat. A lehetséges fenotípusok megjelenésének ezek a korlátjai FEJLŐDÉSI KÉNYSZEREK vagy korlátok (developmental constraints) néven ismertek: „a bias on the production of various phenotypes caused by the structure, character, composition, or dynamics of the developmental system.” (Mayr, 1985).
Főbb kategóriáik: FIZIKAI, SZELEKTÍV (vagy FUNKCIONÁLIS), GENETIKAI, FEJLŐDÉSI KORLÁTOK (vagy KÉNYSZEREK). Tkp. minden korlát a fejlődésre hat, de szokás elkülöníteni szűkebb értelemben vett („sensu stricto”). FEJLŐDÉSI KORLÁTOKAT”.
SZELEKTÍV (vagy FUNKCIONÁLIS) KORLÁTOK: Bizonyos sajátságok azért nem alakulnak ki bizonyos vonalakban, mert mindig előnytelenek, vagy más jelleggel interferálnának.
GENETIKAI KORLÁTOK: Bizonyos fenotípusnál hiányozhat a genetikai variáció. A fejlődési útvonalak különböző mértékben viselik el komponenseik variabilitását, toleranciájuk különbözősége korlátot szabhat a jelleg variabilitásának. Ha a gének új funkciót nyernek az evolúció során, egynél több fejlődési modulban lesz szerepük, ami a változás gátjává válik. A rovarok szegmentpolaritási génjeinek pl. több útvonalban is szerepet játszanak, és (jórészt emiatt) nagyon konzervatívak. Az ilyen pleiotrópia korlátozza az alternatív mechanizmusokat, így nehézkes a változás. A pleiotrópia, a géneknek az a képessége, hogy különböző sejtekben különböző szerepeket játsszanak, egyfajta ellentéte a modularitásnak, összekapcsolja a részeket, önállóságuk ellen hat. Ha két jelleg osztozik egy morfogenetikus útvonalon, az útvonal erősen pleiotropikus lesz, olyan genetikai kölcsönhatások jönnek létre, amelyek erősen korlátozzák a két jelleg szabadságát. A genetikai korlátok, mint a variáció szegényessége, és a genetikai korrelációk a FEJLŐDÉSI KORLÁTOKtól alig elválaszthatók
.
FEJLŐDÉSI KORLÁTOK SENSU STRICTO: Mindössze HÁROMTUCATNYI ÁLLATTÖRZS LÉTEZIK, amelyek az állatvilág valamennyi lényegesebb testtervét reprezentálják. Miért nincsen több?
Hogy ezt megértsük, tisztában kell lennünk azzal, hogy milyen korlátjai vannak a fejlődésnek, mert ezek tükröződnek az evolúció lehetőségeiben. Ide sorolható jelenségek a variáció hiánya vagy szegényessége, ideértve a morfogenetikai kapacitást is (a struktúra kifejlődéséhez szükséges sejtek, gének, fehérjék hiánya), és a jellegek közötti erős korrelációk, amelyek eredhetnek a szövetek kölcsönhatásaiból, vagy a gének pleiotiotrópiájából. A morfogenetikus konstrukciós szabályok is korlátozzák a lehetséges fenotípusok számát (MORFOGENETIKAI KORLÁTOK a FEJLŐDÉSI KORLÁTOKON BELÜL).
Már Bateson 1894-ben megállapította, hogy egy organizmus fejlődése csak néhányféle módon változhat meg, térhet el a normálistól. Pl. Az elmúlt 300 millió évben sokféle gerinces végtag alakult ki, de pl. olyan, amelynek a középső ujja rövidebb a többinél, soha. A természetes populációkban is csak minimális számú módja van egy végtag megváltozásának. A hosszabb végtag kialakulásánál pl. a humerus hosszabbodik meg általában, de sosem lesz pl. két humerus egymáshoz csatlakozva. Valószínűleg a végtag fejlődését a reakció-diffúziós modell szerint képzelhetjük el, ami korlátozza a kialakulás módozatait.
A lehetséges fenotípusok számát és megjelenését korlátozzák azok a kölcsönhatások, amelyek
-
-
a molekulák
-
a modulok
-
között lehetségesek
Fizikai korlátok:
-
Fizikai törvényekből erednek: a diffúzió, a hidraulika, a gravitáció, a mechanika törvényei az élőlényekre ugyanúgy érvényesek, így csak bizonyos fenotípusok jelenhetnek meg, nincsen pl. 8 méter magas légy, vagy 1000 tonnás szárazföldi állat.
-
A szövetek elaszticitása és szakítószilárdsága is korlátozó tényező.
-
A Drosophila 2 mm hosszú spermiumához speciális tubulin kell!
-
A morfogenezis hat alapvető sejt-viselkedési formájának (osztódás, növekedés, formaváltoztatás, vándorlás, sejthalál, mátrix szekréció) megvannak a maga fizikai korlátjai, így korlátozott, hogy milyen állati struktúrák alakulhatnak ki
-
A különféle szövetek közötti interakciók során a sejtrétegek, csöves képletek, pálcaszerű képletek csak korlátozott módon léphetnek kapcsolatba
Morfogenetikai korlátok
-
A morfogenetikus konstrukciós szabályok is korlátozzák a lehetséges fenotípusok számát
-
A természetes populációkban is csak minimális számú módja van egy végtag megváltozásának
-
A fogaknál hasonló elvek érvényesülnek
-
Bizonyíték a reakció-diffúziós modell mellett: kísérletes embriológia, összehasonlító anatómia, sejtbiológia területéről.
-
Ha axolotl végtagbimbóját colchicinnel kezelik, a sejtek száma lecsökken a végtagbimbóban. Nemcsak az ujjak száma lesz majd kisebb, hanem megjósolható, mely ujjak, milyen sorrendben tűnnek el. A jóslás alapja: a „tiltott” morfológiák, és a matematikai modellek. Amilyen szalamandra végtagok kísérletesen kifejlődtek, azokhoz hasonló fajok is vannak a természetben!!!
Filetikus korlátok
-
Az új struktúrák kialakulását korlátozó filetikus kényszerek az élőlény evolúciójából következnek, és a fejlődésének genetikai hátteréből következnek.
-
Ha egy struktúra induktív interakciókkal jön létre, nehezen elképzelhető, hogy fejlődése „a nulláról” újraindulhat
-
A notochord pl., amely funkcionális Amphioxusban, degenerálódik a felnőtt gerincesekben. De!!!! Tranziens jelenlétére mégis szükség van, mert specifikálja a velőcsövet
-
Hasonlóan: Waddington 1938-ban rájött, hogy a csirke elővese vezetékét „csökevényes” jelzővel illetik, mivel nem vesz részt a vizelet koncentrálásában, részét képezi a húgycsőbimbó, amely a funkcionális vese kialakulását indukálja
-
A filetikus korlátok egyik legszebb példája: az erszényesek végtagjai alig-alig mutatnak változatosságot! Az embriónak már nagyon korán ki kell fejlesztenie erőteljes végtagjait, hogy megragadva anyja szőrét fel tudjon mászni az erszénybe. Gyakorlatilag megszűnt a változatosság!
-
Ha a gének új funkciót nyernek az evolúció során, egynél több fejlődési modulban lesz szerepük, ami a változás gátjává válik.
-
A rovarok szegmentpolaritási génjei pl. több útvonalban is szerepet játszanak, és nagyon konzervatívak. Az ilyen pleiotrópia korlátozza az alternatív mechanizmusokat, így nehézkes a változás (ld. előbb is).
-
Az emlősök fejlődési korlátjainak alapjainál Galis szerint az emlősöknek azért van csak 7 cervikális (nyak) csigolyájuk, mert az őket specifikáló Hox-gének az emlősökben összekapcsolódtak a sejtosztódással. A Hox-gének kifejeződésének változásai így a sejtosztódást is megzavarnák, ami rákhoz vezetne. Mellette szól, hogy rendkívül erős a szelekció a nyakcsigolyák számának változása ellen embernél!!!!
-
Egészen a legutóbbi időkig úgy gondolták, hogy a fejlődés első időszakát a legnehezebb megváltoztatni - vagy teljesen megzavarja az embrió fejlődését, vagy radikálisan új fenotípushoz vezet. Nem így van… a lárvák egy kicsit megváltozhatnak a Molluscaknál, ha a citoplazmás determinánsokat megváltoztatják, de ugyanúgy Mollusca alakul ki! A tengeri uborkáknál a citoplazmás determinánsok eloszlásának megváltoztatása akár lárvastádium nélküli fejlődéshez vezethet, de ugyanolyan állat fejlődik ki!
-
Tehát a korai fejlődés plasztikus — a késői nyilvánvalóan az.
-
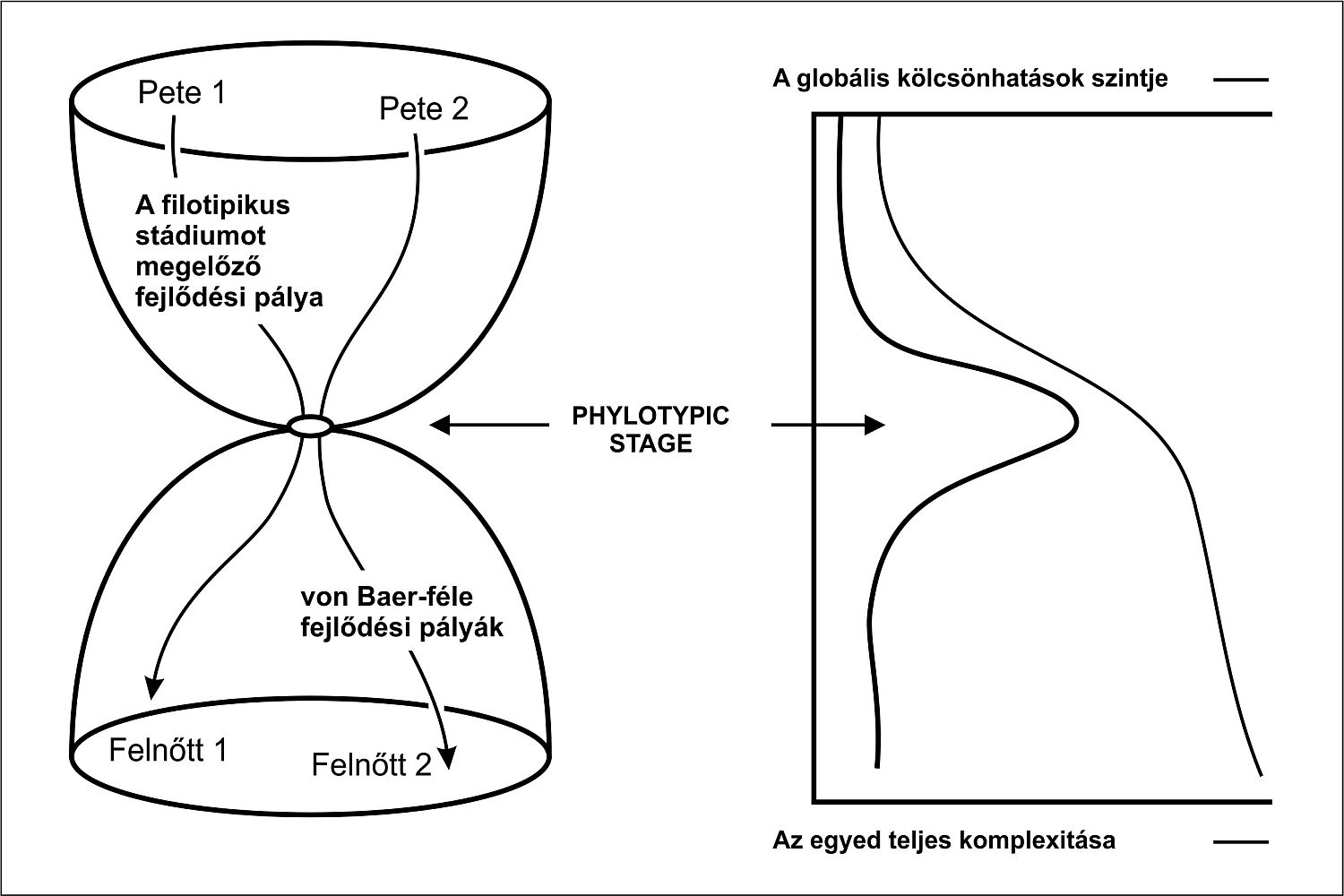
A FILOTIPIKUS SZAKASZ, A „KÖZÉPSŐ” SZAKASZ MEREV (6. ábra).
-
Előtte-globális kölcsönhatások, nagyfokú reguláció
-
Utána sok induktív esemény - de modulon belül.
-
A pharyngula stádiumban a modulok kölcsönhatásban vannak!- emberi többszörös fejlődési rendellenességek is emellett szólnak - ez a legérzékenyebb fejlődési szakasz
-
HA EGY ÉLŐLÉNY EGYSZER GERINCESSÉ VÁLIK, VALÓSZÍNŰLEG LEHETETLEN MÁSSÁ ÁTALAKULNIA
13.1. ábra - A fejlődés homokóra-modellje: a modell lényege, hogy az élőlény fejlődési moduljai között a kölcsönhatás szintje a filotipikus stádiumban maximális, előtte és utána összehasonlíthatatlanul nagyobb a függetlenségük.
 |