Darwin, mint sokszor említettük, úgy gondolta, hogy az evolúció apró lépésekben, fokozatosan halad, ez a megközelítés a gradualizmus. A fajok eredetében kifejtésre kerül, hogy még a legbonyolultabb szerv is apró módosulásokon keresztül jöhetett csak létre, mert ellenkező esetben sutba kéne vágni az elméletet… Thomas Henry Huxley, „Darwin bulldogja” hívta fel a figyelmet arra, hogy a darwini elmélet akkor sincsen semmi baj az elmélettel, ha az evolúció „ugrásokkal” halad. Jóval későbbi evolúciós biológusok hasonló nézeteket vallottak, köztük a paleontológus Schindewolf, aki szerint: a magasabb taxonok közötti különbségek „diszkontinuusan, szaltációval alakultak ki” (a szaltáció szó a latin saltus szó származéka, ugrást jelent). Híres mondása: „az első madár egy hüllő tojásából bújt ki”.
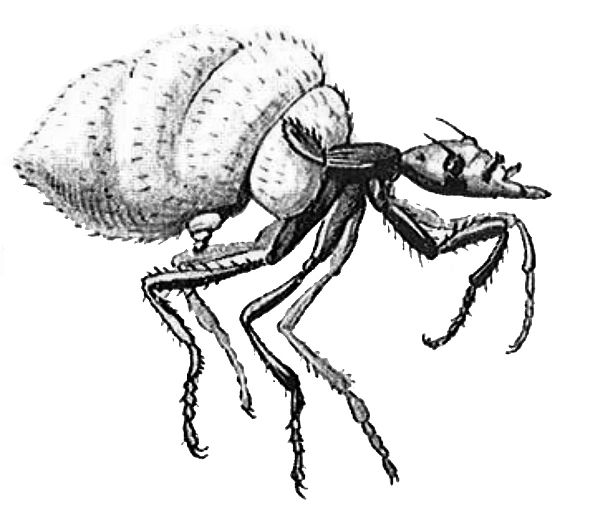
Richard Goldschmidt, a nagyszerű genetikus, aki az elsők között javasolta, hogy a gének anyagcsere utakat és fejlődési útvonalakat szabályoznak The Material Basis of Evolution (1940) című művében amellett érvel, hogy a fajok, és a magasabb taxonok nem a fajokban meglévő genetikai variációra gyakorolt szelekció eredményeként jönnek létre, hanem „egyetlen evolúciós lépésben, mint teljesen új genetikai rendszerek”. Posztulálta, hogy a kromoszómák anyagának komolyabb megváltozása, vagy „rendszermutációk” („systemic mutations”) drasztikusan megváltozott élőlényekhez vezetnek, melyeknek legtöbbjének alig van esélye a túlélésre, de néhány „reményteljes szörnyeteg” új életformákhoz alkalmazkodhat. Példája a Drosophila melanogaster billérjeit részlegesen szárnnyá transzformáló mutáció volt. A mutánsok haltérái hasonlítottak a szintén Diptera légy, a termeszvárban élő Termitoxenia szárnyához 8. ábra.
Amellett, hogy sok példa van a finom átmenetekre a jelenlegi és a régvolt élővilágban egyaránt, az is vitathatatlan, hogy nyilvánvaló „rések” vannak, különösen a magasabb taxonokba elkülönített élőlények között. Jelenleg nem él átmeneti forma pl. a cetek és a szárazföldi emlősök között. A fenotipikus résekre pontosan a ceteknél találjuk a legegyszerűbb magyarázatot. Itt voltak átmeneti formák, csak kipusztultak. Az is könnyen belátható, hogy két, (akár nagyon) különböző forma közötti „átmeneti lény” nem feltétlenül mutat átmeneti jegyeket, mivel a két utód nagyon eltérő irányba fejlődhetett. Klasszikussá nemesedő példa, hogy a ma élő állatok közül a vízilovakkal állnak a legközelebbi rokonságban, de a korai fosszilis ceteknek a külseje a legcsekélyebb mértékben sem hasonlított a vízilovakéhoz. A fosszilis anyagban vannak példák a magasabb taxonok graduális evolúciójára, de számos magasabb taxon semmilyen átmeneti formával nem rendelkezik.
Az evolúcióbiológia leghosszasabb vitája, hogy a rések a fenotípusok között a fosszilis anyag hiányosságainak köszönhetők, az átmeneti formák léteztek, csak nem leltek rájuk, vagy az evolúció ténylegesen haladhat szaltációval. A szaltáció egyetlen, nagy hatású mutáció fixálódásával jönne létre. Érdemes végiggondolni, hogy ez a koncepció teljesen mást jelent, mint a szaggatott egyensúly. A szaggatott egyensúlynál a morfológiai változás lehet (de nem szükségszerűen) folyamatos, átmehet sok átmeneti formán, de olyan gyorsan, és földrajzi szempontból annyira lokalizáltan, hogy a fosszilis anyagban nem folyamatos (=diszkontinuus) változás tükröződik. A szaltációs hipotézis szerint az átmeneti formák sohasem léteztek—a mutáns élőlények drasztikusan különböztek szüleiktől.
A gradualizmus-szaltáció kérdésben való döntéshez különbséget kell tennünk a taxonok és azok jellegei között. A magasabb taxonok sokszor különböznek egész sor jellegben, pl. a madarakat számos jelleg különíti el a dinoszauruszoktól. A gradualisták szerint a magasabb taxonok sok jellege függetlenül és egymást követően fejlődtek ki, azaz mozaik evolúció történt. A mozaik evolúcióra sok példa ismert a mai és a régmúlt korok élőlényeinek köréből, vitatott azonban, hogy a magasabb taxonok megkülönböztető jellegei folyamatosan, vagy ugrásszerűen fejlődtek ki, pl. a madarak farokcsigolyáinak redukciója és összeolvadása hogyan zajlott.