Bármely jelleg eredetének és evolúciójának megértéséhez szükséges a filogenetikai viszonyok ismerete. Általában a filogenetikai viszonyokat olyan közös jellegekből következtetik ki, amelyek származtatottak, így viszonylag közeli rokonságot jeleznek. Sokszor nehéz eldönteni, melyek legyenek ezek a jellegek.
Még tíz éve is feloldhatatlannak tűnt a gerinctelen csoportok rokonsági viszonya. A morfológia alapján csaknem lehetetlen feladat a törzsek rokonsági viszonyainak tisztázása! Egy vonzó, és az utóbbi évtizedekben magától értetődően adódó lehetőség a morfológia megkerülésére: molekuláris genetikai jellegek felhasználása az állati filogenezis vizsgálatára, a DNS divergenciája alkalmas a rokonsági viszonyok vizsgálatára.
Az utat a DNS filogenetikai felhasználásának kifinomulása, a molekuláris és bioinformatikai módszerek javulása, a konzervált szabályozó génekre vonatkozó új adatok, a régebben különböző eredetűnek vélt struktúrák rokonságára vonatkozó bizonyítékok és az előítéletmentes és félelmetes adatmennyiséget feldolgozó programok nyitották meg.
Felhasználhatónak bizonyult továbbá a rokonsági viszonyok finomításához, hogy bizonyos gének egyes vonalakban hiányozhatnak vagy kapcsoltsági viszonyaik eltérőek lehetnek. A szekvencia-genetikai alapú módszerek kombinálhatók a morfológiával is.
Mindezek alapján, a közös morfológiai, fejlődési és genetikai jellegek alapján az állatvilág felosztható (13. ábra):
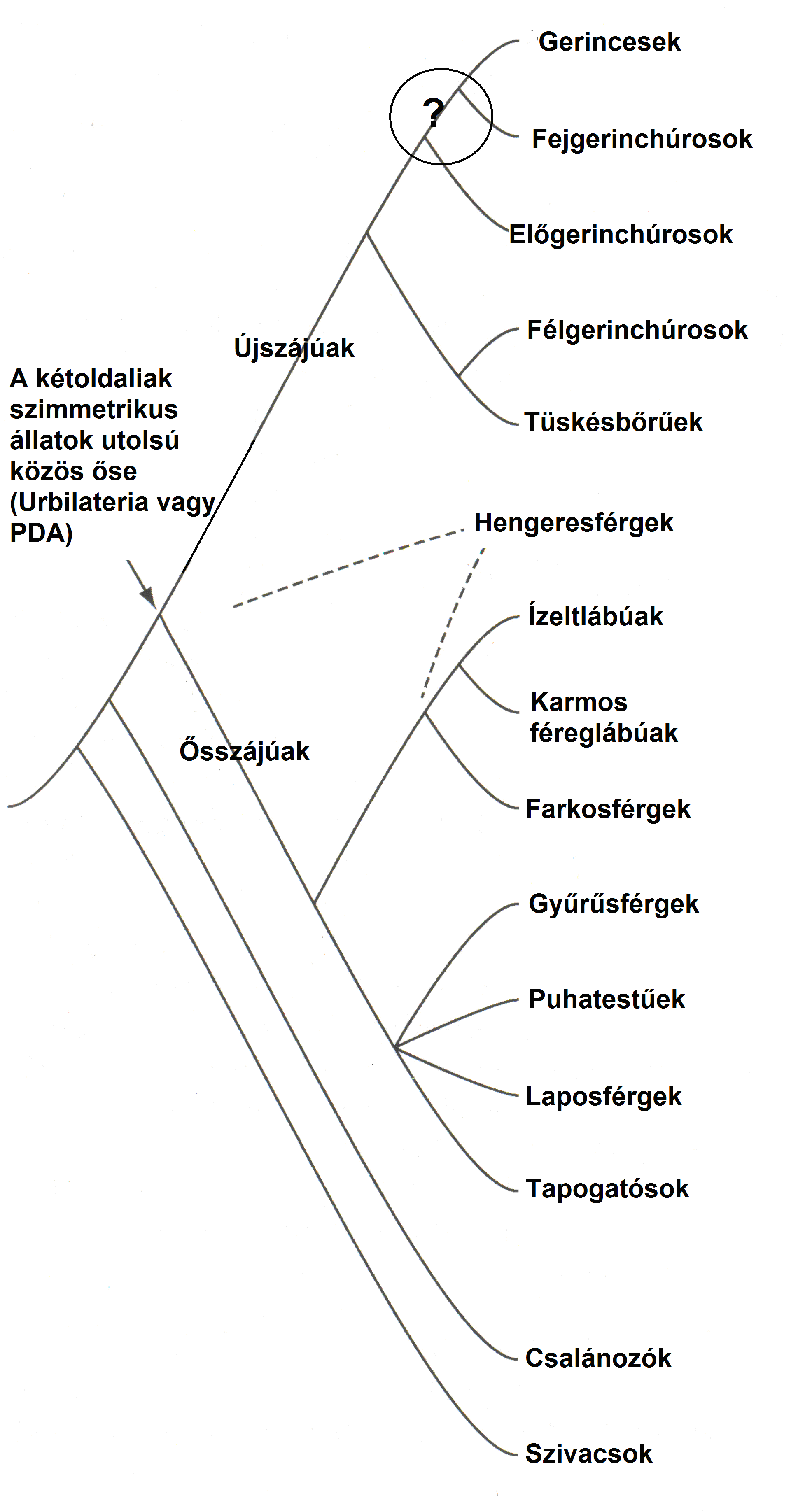
4.13. ábra - Közismert törzsek rokonsági viszonyai. Kiemeljük, hogy a gerincesek legközelebbi testvércsoportjának újabban az előgerinchúrosokat tartják, a hengeresférgek besorolása pedig változatlanul problematikus (szaggatott vonal).
 |
(I) Porifera- szivacsok, „álszövetesek”
(II) Cnidaria és Ctenophora—csalánozók és bordásmedúzák: radiális szimmetriájú diploblasztok, legfeljebb minimális mezodermával.
(III) Bilateralia –triploblasztok, kétoldali szimmetriával. Alapvetően feloszthatók ősszájúakra és újszájúak
Ősszájúak: Ecdysozoa (exosceleton - növekedés, vedlés) – monofiletikusak valamint Lophotrochozoa (puha szöveteik legnagyobb része kapcsolatban van a külvilággal, csillókat használnak evéshez és mozgáshoz) - monofiletikusak
Újszájúak: Echinodermata+ Chordata: száj másodlagosan alakul ki, a blasztopórusból (ősbélnyílás a gasztrulában) lesz az anus.
Fontos további, nem várt eredmények:
- Arthropodák és Annelidák külön kládba tartoznak
- laposférgek puhatestűekkel - bár nem közeli - rokonok, Lophotrochozoák közé sorolhatók
- hengeresférgek helyzete bizonytalan, de Ecdysozoák
- tehát a cölómanélküli (valódi testüreg nélküli) Nematodák és a laposférgek biztosan nem „őseink” azaz fejlődési vonalunkban nem voltak ilyen élőlények, sőt, nagyon távoli rokonok maguk is
A modern állatvilág kialakulásában kulcsváltozások történtek a kambriumban (545-490 millió éve). Darwin óta tudott, hogy ekkor valami történt - az állati változatosság drámai mértékben növekedett. A legtöbb modern törzs első megjelenése ide köthető (egészen az 1940-es évekig a kambriumból kerültek elő a legrégebbi fosszíliák).
A NAGY, KOMPLEX ÁLLATOK MEGJELENÉSÉT ÉS RADIÁCIÓJÁT A KORAI-KÖZÉPSŐ-KAMBRIUM 10-25 MILLIÓ ÉVE ALATT „KAMBRIUMI ROBBANÁSKÉNT” SZOKÁS EMLÍTENI.
Több okból feltehető, hogy a legtöbb modern törzs már előbb megjelent:
Némelyeknek már fosszíliáikat is találtak (csalánozók, szivacsok, puhatestűek(?))
Nyomfosszíliák (trace fossils) vannak, amelyek triploblasztikus Bilateráliákra utalnak
Ediacara fauna - sikertelen kísérlet? - azért nem hasonlítanak, mert ősiek?
Molekuláris óra (riboszomális RNS és „szuperkonzervatív”fehérjék alapján): a legtöbb törzs 650- >1000 millió éve jelent meg (ld. Molekuláris evolúciós ráták).
A primitív bilateráliák talán túl kicsik voltak (kevés oxigén miatt?). Kihalás lehetett a proterozoikum-kambrium határon - talán ez nyitotta meg a niche-ket – vetélkedésük talán kedvezett a külső vázas formák kialakulásának, ragadozók kialakulásának, egyáltalán az INNOVÁCIÓKNAK.
Testtervnek nevezzük az egy törzsbe tartozó élőlényekre jellemző genetikai és fejlődési mintázatot, amely rögzíti az élőlények testének alapvető geometriai mintázatát olyan módon, hogy meghatározza az anatómiai részek egymáshoz viszonyított helyzetét, ezáltal megszabja az élőlényre jellemző összes alaki sajátságot. Tulajdonképpen egy kaszkádot indít el, aminek végeredménye az élőlény. Alapvetően törzsekre vonatkozó fogalom, de minden törzs alatti rendszertani szinten értelmezhető. Feladata az egyes anatómiai részek pozícionális rögzítése. Ide kapcsolódó fogalom az archetípus: monofiletikusan származtatható élőlényekre jellemző legegyszerűbb morfológiai, anatómiai és genetika mintázatok összességét értjük rajta. Jó közelítésben azonosítható a – leegyszerűsített – testtervvel.
Jelenleg kb. 35 állattörzs létezik különböző alaptesttervekkel, melyek mindegyike a kambriumban jelen volt! Azóta nincs új alaptestterv - de a testtervek "kidolgozása" új élőlénytípusokhoz vezetett
Multicelluláris (soksejtes) élet
Új, több mint 2000 millió éves, egyelőre problematikus besorolású, soksejtű lényeket találtak 2010-ben. Legkorábbi multicelluláris (Metazoa) élőlények 1100-1200 millió évesek, és ma is besorolható szivacsoktól származnak (velük együtt mászásnyomok is előkerültek, egyelőre nem tudni, milyen élőlényektől).
Főként miért bizonytalan az állati testtervek kialakulásának vizsgálata?
Nagy rések a fosszilis emlékekben
Óriási különbségek a radiálisan szimmetrikus állatok és a Bilateráliák anatómiájában, fejlődésében, genomjának sajátságaiban
A bilateráliák korai történetét sűrű homály fedi
A paleontológusok bevezették a diszparitás (disparity) fogalmát a testtervek közötti különbségek jellemzésére, a diverzitás fogalmát az adott alapvető testtervű állatok közötti különbségek jellemzésére. Bizonyos testterv diverzifikációja jóval könnyebben megközelíthető, a különféle testtervek eredete egyelőre nem.
Az emlősök kialakulása a hüllőkből
Amikor az emlősök kifejlődtek a hüllőkből, nagyfokú változásoknak kellett létrejönniük számos jellegben. A folyamatban makroevolúciós változás zajlott le – alapvető kérdés: hogyan zajlott le a tranzíció (átmenet)?
A legkorábbi emlős fosszíliák között vannak a Morganucodontidok, pl. Kuehneotherium a triász-jura határon 200 millió évvel ezelőttről (ma az emlősök eredetét kb. 230 millió évvel ezelőttre teszik). Nem tudjuk, hogy elevenszülő volt-e vagy laktált? Közvetlenül nem tudni, de emlős állkapcsa, tartása, fogstruktúrája volt. Melegvérűségét is sok minden valószínűsíti.
A Morganucoidontidok elődei, az emlősszerű hüllők (Synapsidák) ~100 millió év alatt fejlődtek a triász végéig, az első igazi emlősökig. Bizonyos Synapsidák még a jurában is éltek – a dinoszauruszok már javában virágzottak.
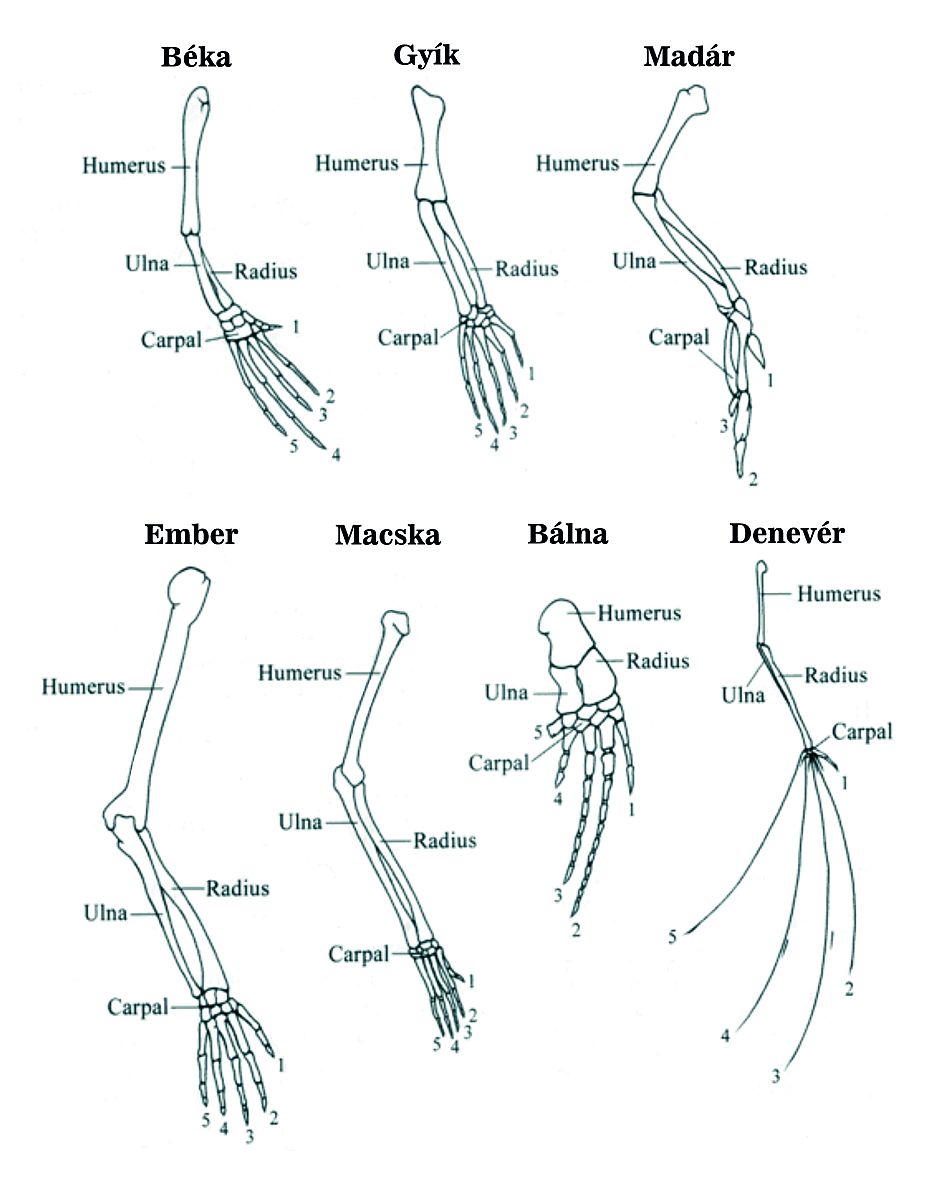
A négylábúak között a fosszíliákból legjobban rekonstruálható jellegek a mozgással és táplálkozással – egyszerűen a csontokkal és fogakkal - kapcsolatosak. A hüllők fogai viszonylag differenciálatlanok és egyszerűek. Állkapcsuk alsó és felső része hátul ízesül ahol egyszerű “csukó” izmaik vannak. Az emlősök állkapcsához pofaizmok tapadnak – ezek körülveszik a fogakat és erőteljesebb, pontosabb a záródás. Az evolúció során az állkapocs ízesülése előre mozdult, a hátsó rész csontjai “felszabadultak” és a belső fül csontjaivá transzformálódtak.
Az emlősök evolúciójának 3 fő szakasza 15. ábra:

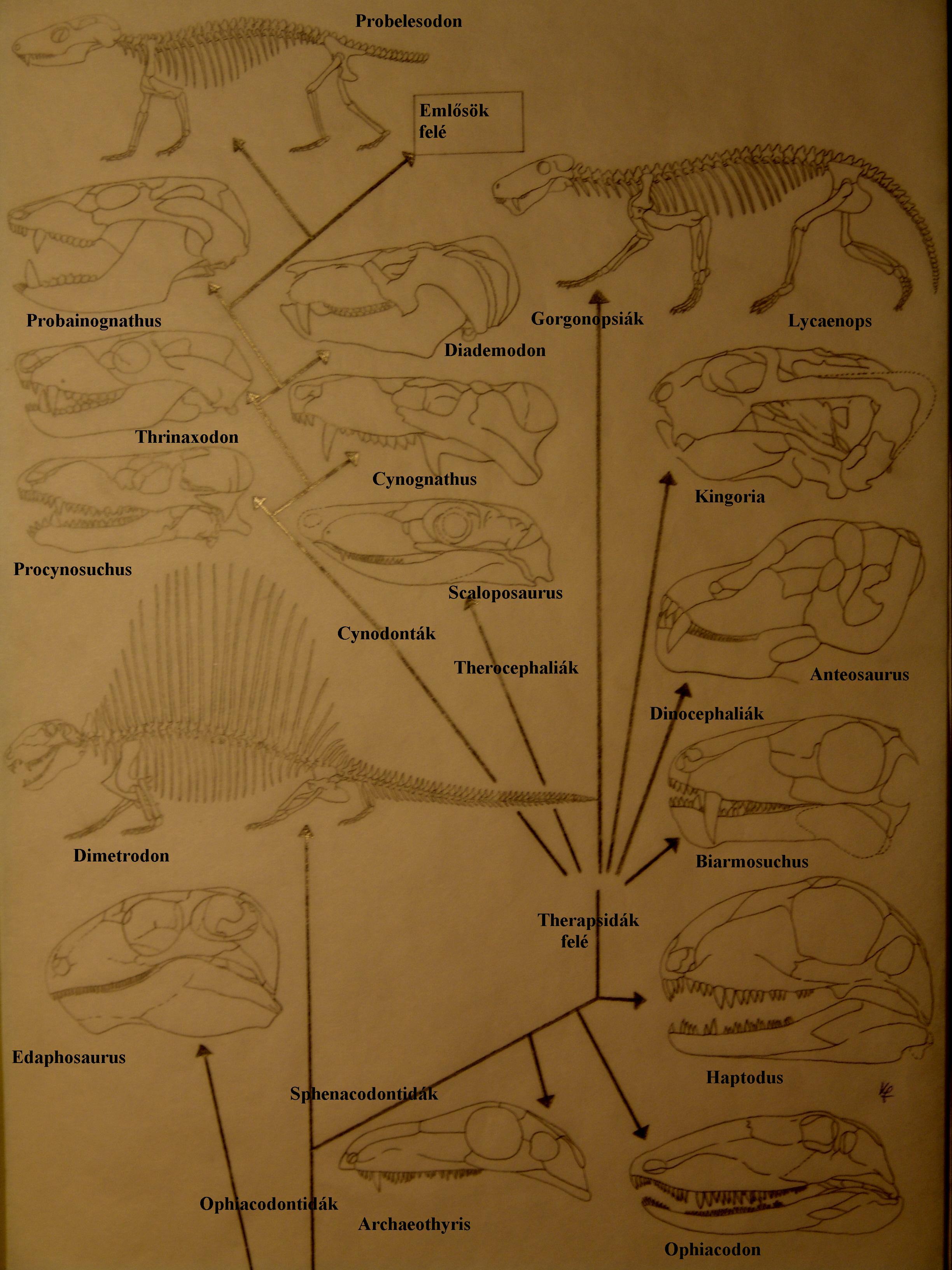
1. Pelycosaurusok: pennsylvaniai karbon és perm rétegekben az USA DNy-i részén találják meg az Archaeothyrist (14. ábra) – 300 millió éve élt, gyíkszerű, 50 cm hosszúságú, az emlősök távoli őséhez nagyon közel álló forma. Fontos anatómiai jegyük egy nyílás a szem mögött (temporal fenestra), amelyen izom haladt át. Az izom az állkapocs zárására szolgált és az erős állkapocszáródás első jeleként értékelhetjük ebben a vonalban. A temporális fenestra a Synapsidák meghatározó jellege. Pelycosaurusok: fogdifferenciáció csekély, csúszómászó tartás, 260 millió éve nagyobbrészt kipusztultak.
2. A Pelycosaurusok Sphenacodontida csoportjából fejlődtek ki a Therapsidák (Dél-Afrika, Karroo medence leletei) a késői permben – korai triászban. Temporális fenestrájuk nagyobb és emlősszerűbb, fogaik differenciáltak. A későbbi formáknál másodlagos szájpadlás alakul ki, lehetővé téve, hogy egyszerre egyenek és lélegezzenek.
3. Az aktívabb, melegvérű Therapsidák egy csoportja a Cynodonták, melyek állkapcsa hasonlít a modern emlősökére. Fogaik multicuspidálisak (többcsúcsúak) és differenciáltak. Állkapcsuk ízesülése esetén az az elv érvényesül, hogy az evolúcióban az 1. struktúra 2. struktúra átmenetnél nem lehet köztes funkcióképtelen állapot! Az A struktúrából lesz A+B, abból B.
A Cynodonták egyik vonalából alakultak ki a modern emlősök, pl. a Kuehneotherium.
A teljesen pontos Cynodonta vonal nem ismert, de a Probainognathus közel van a hiányzó láncszemhez. A fosszílis rekord minden más ilyen átmenetnél jobban dokumentálja az emlősök eredetét – jól lehet tanulmányozni a nagy átmenetek lefolyását. Következtetések:
a hüllő - emlős jellegek fokozatos átmenetet mutatnak. Nagyskálájú változások jöttek létre adaptációval, a változások biztosan adaptívak és a természetes szelekció által jönnek létre. Az általános evolúciós modell, ami mellett az emlősszerű hüllők evolúciója szól: a természetes szelekció kumulatív módon hat egy hosszú időperiódus (adott esetben legalább 40 millió év) alatt, sok kis skálájú változás hatására a hüllőkből emlősök alakulnak ki.
Az extrapolatív teória szerint a makroevolúciót ugyanazon ágensek hozzák létre, mint a mikroevolúciót, azaz a természetes szelekció hozza létre az adaptív javításokat, akárcsak a fajon belül és speciációkor, csak sokkal nagyobb időskálán.
Az extrapolatív modell nem az egyetlen modell a nagyobb csoportok kialakulására, de mindmáig a legfontosabb és fosszíliákkal leginkább alátámasztott. Tulajdonképpen ezt értjük “neo-darwini makroevolúciós elmélet” alatt.
A populációs genetikában a neo-darwinizmus a mikroevolúciós változásokat a meglévő variánsok frekvenciaváltozásaival magyarázza. A természetes szelekció a fajok - molekuláris és klasszikus genetikai vizsgálata során feltárt - „szokásos” variabilitási szintjén hatva elégséges mindazon változások megmagyarázásához, melyeket ismerünk!
Nem kizárt, hogy más, akár nagyobb csoportok nem adaptív, hanem azonnali változással is kifejlődhettek, bár egyértelmű példát erre nem ismerünk. Fontosnak tartjuk megjegyezni, hogy a hüllő ® emlős átmenet jelentős leegyszerűsítése volna azt gondolni, hogy ez egy egyenes vonalú fejlődés. Minden jelentősebb stádiumban adaptív radiáció figyelhető meg!
Az evolúció igen ritkán generál új anatómiai struktúrákat "a semmiből" - egy létező struktúra módosul - az evolúció toldás-foldás, barkácsolás (angolul „tinkering”).
Pl. az emlősök középfülében 3 csont található, amelyek a dobhártyáról viszik át a hangrezgést a belső fülre. Hüllőkben a koponya és az állkapocs ízesülése os quadratum és os articularis (alsó álkapocs része) között jön létre (16. ábra).
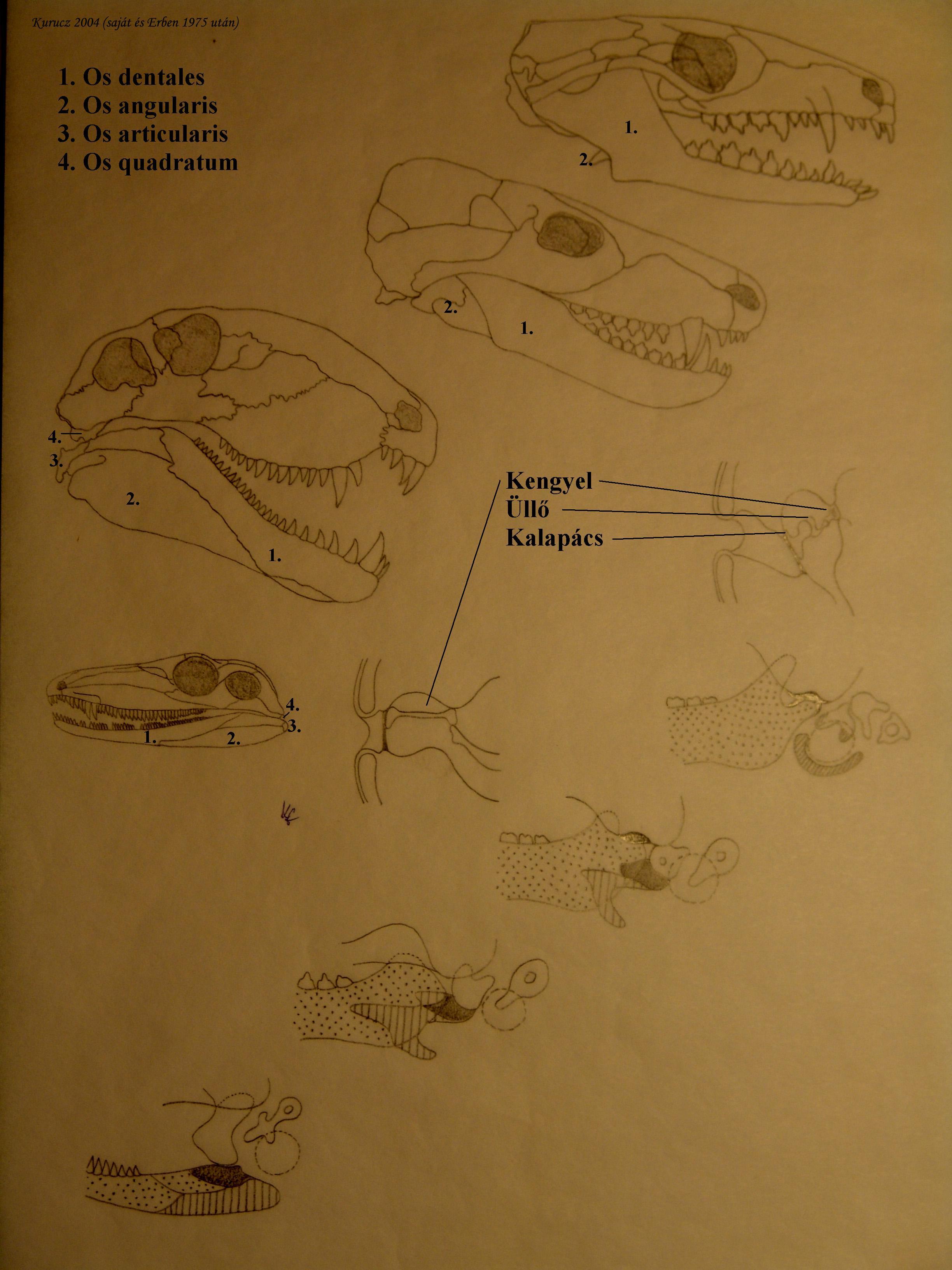
Emlősök: az alsó állkapocs 5 csontjának szerepét 1 csont (os dentales) veszi át.
Az articularisból kalapács, a quadratusból üllő alakul ki.
Másik példa: gerinces vese .
Madaraknál, emlősöknél az egyedfejlődés során a pronephros (átmeneti) - mesonephros - vese átmenet zajlik le.
A mesonephrosból jönnek létre a gonádok: testis és az ovárium szomatikus sejtjei.
Halaknál, kétéltűeknél a lárvában a pronephros funkcionál, a felnőtt egyedben a mesonephros funkcionál.
Az "ősök" veséje embrionális szerv, de módosult, és a gonádok fejlődéséhez nélkülözhetetlen struktúrát hoz létre.
Az evolúciós fejlődésbiológia mutatta meg, hogy a makroevolúciós változásokat két alapelv teszi lehetővé:
-
a modularitás
-
és a molekuláris parszimónia
Az állati testszerveződés és diverzitás általános sajátsága a modularitás (az állatok teste kisebb, viszonylag független egységekből áll, amelyek egyben a fejlődésük egységei is).
-
Alapkérdés, hogyan változhat meg egyáltalán egy embrió fejlődése, ha a fejlődés annyira finoman össze van hangolva, és annyira komplex anélkül, hogy anélkül felborulna az egész rendszer?
-
A változások azért lehetségesek, mert a fejlődés diszkrét és kölcsönhatásban lévő modulok sorában zajlik! Modul lehet egy végtag, egy szelvény, a szárny egy része st.
-
A (nagyobb) bilateráliák ismétlődő egységekből, vagy modulokból állnak: különböző törzsek szelvényei
-
Gerincesek csigolyái és kapcsolódó struktúrái az alapegységei a testterv szerveződésének. Hasonlóan: sok testrész maga is ismétlődő struktúrákból áll: négylábúak lába, rovarok szárnya.
-
De modulok a morfogenetikus mezők, jelátviteli utak, imaginális diszkuszok, sejtvonalak, paraszegmentek, szervkezdemények is!
-
Most egy ideig szorítkozzunk a morfológiai értelemben vett modulokra. Már a korai kísérleti embriológusok számára is világos volt, hogy egyes modulok elkülönülten fejlődhetnek más moduloktól. Twitty a harmincas években szalamandralárva végtagbimbóját és szemmezőjét ültette át. (Egyik) legfontosabb trend az állati fejlődésben: a moduláris elemek kiegyénülése (individualizációja).
-
Az Onychophora szelvényei viszonylag egységesek <=> az Arthropoda szelvényei kiegyénültek
-
Az emlősök fogazata kiegyénült fogakból áll <=> A hüllők fogazata viszonylag egységes
-
Az ilyen különbségek meghatározóak=> megértésük a taxonok evolúciójának elemzéséhez nélkülözhetetlen.
Bateson szerveződési szintjei
Bateson: szerveződési szinteket ír le, Materials for the Study of Variation (1894) című művében elsőként vesz fel darwini perspektívát a különféle testszerveződések evolúciójával kapcsolatban. A részek ismétlődésére koncentrál: lajstromba szed nagyszámú ritka, de a természetben előforduló variánst, amelyek különböztek a normális egyedektől akár ismétlődő elemeik számában, akár kiegyénültségében. Szerinte ezek a fajon belüli variációk rávilágíthatnak a nagyobb skálán folyó változásokra. Ilyen változások bizonyos frekvenciával fellépnek a populáción belül - plauzibilis, hogy a fajok diverzifikálódásában is lehet szerepük.
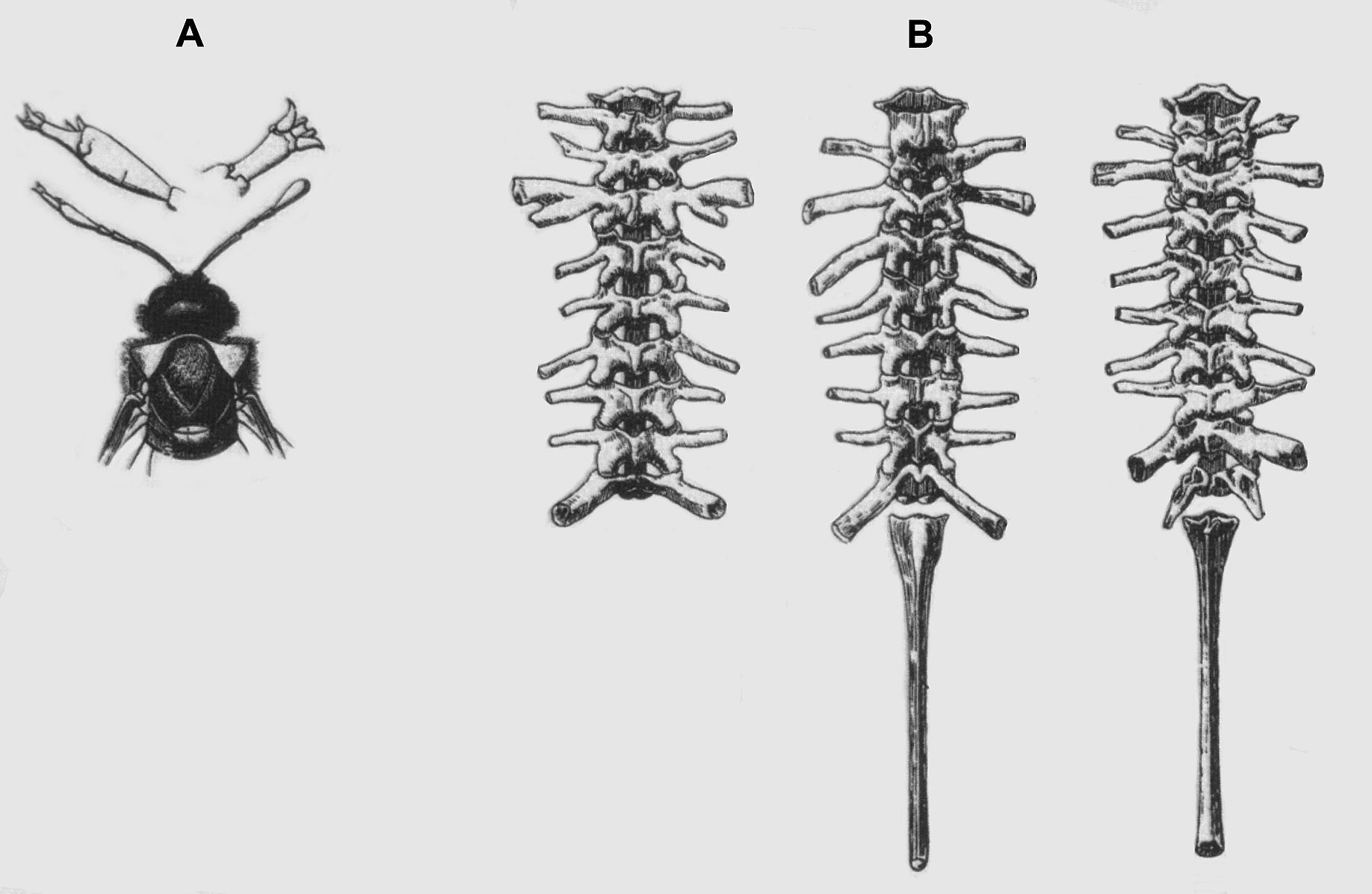
Kimondottan fontosak az evolúcióban - Bateson organizációs elvei és a vizsgált variánsok (18. ábra) sokat segítettek az állati test moduláris természetének alapját képező genetikai és fejlődési folyamatok logikájának megértésében - kulcsfontosságú gének, a homeotikus gének felfedezése is folyománya munkájának.
4.17. ábra - Homeotikus mutáció: csáp transzformációja lábbá; különböző csigolyák transzformációi gerinceseknél egy XIX. századi ábrán
 |
A moduláris felépítésű állatoknál 5 alapvető, evolúciós szintű morfológiai változás biztosan fontos az adaptáció szempontjából:
-
1. Változások az ismétlődő részek számában. Bateson az ilyen jellegű, a fajon belüli különbségekre alkalmazta a meristic variation kifejezést, pl. bordák száma (19. ábra)(19. ábra).
4.18. ábra - Ismétlődő részek különböző állatcsoportokban. Az elemek számában fajon belül és fajok között is variabilitás tapasztalható (meristic variation)

-
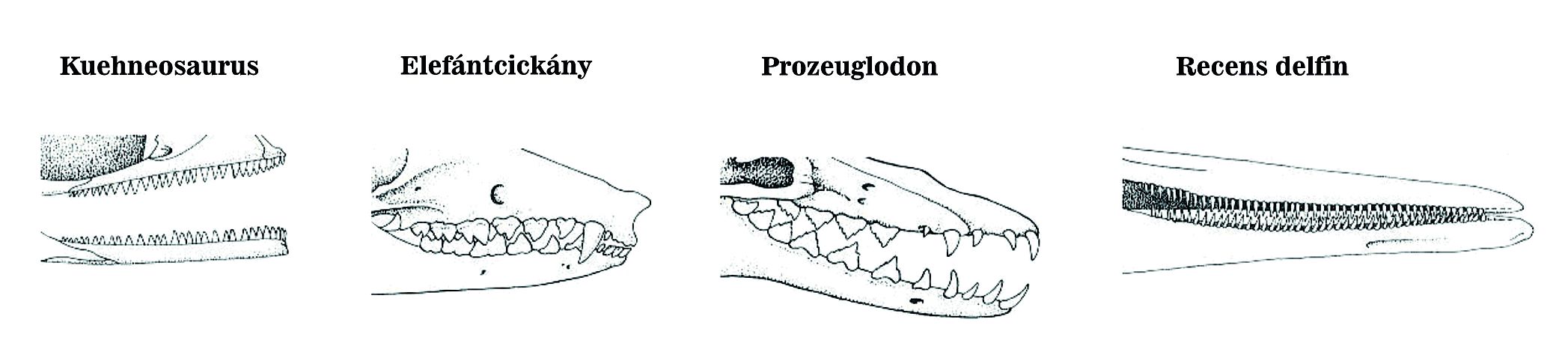
2. A sorozatszerűen homológ (serially homologous) részek kiegyénülése (ide tartoznak pl.: rovarok lábai, a gerincesek csigolyái) (20. ábra). Nagyon ritkán előfordul, hogy a kiegyénülés elvész, szép példa erre a fogascetek fogsora (21. ábra).
4.20. ábra - A fogazat kiegyénülése ritka esetben visszafordulhat. A hüllőknél a fogazat egységes, az emlősöknél (elefántcickány) heterodont fogazatot találunk, azaz a fogak egyediek, a kezdetleges bálnáknál már megfigyelhető a fogak dedifferenciációja (Prozeuglodon), a modern fogascetek (delfinek) fogazata homodont, azaz a fogak egységesek
-
3. A különböző vonalak homológ részeinek diverzifikálódása. 22. ábra A homológ részek morfológiai diverzifikálódása a különböző vonalak között az állati evolúció egyik legfontosabb trendje. Két struktúrát akkor nevezünk homológnak, ha közös történetük van, függetlenül a funkciójuktól.
-
4. Az evolúció alapvető mintázataihoz sorolhatók az evolúciós innovációk (újítások). Evolúciós újítások - kifejlődhetnek egy már létező struktúrából, vagy de novo (újonnan). Új célokhoz adaptálódnak – külön fejezetet szentelünk nekik, itt csak alapproblémaként kerülnek említésre.
Az olyan „új sajátságokat” mint a madártollak, vagy a szárazföldi négylábúk lábát – amelyek elősegítik az élőlények alkalmazkodását egy speciális életformához - evolúciós újításoknak (evolutionary innovations) nevezzük.
Hogyan alakulnak ki? Az erről szóló vita Darwin óta tart és máig nem hagyott alább.
Az élőlények mérete és formai lehetőségei korlátlanoknak tűnnek, de ha jobban megnézzük az igazán új struktúrák száma kimondottan csekély. Elképzelhetünk pl. „angyali” lényeket a vállukból kinövő szárnyakkal, de a valóságban a gerincesek szárnyai mindig módosult mellső lábak. A modern emlősök igen változatosak, de minden struktúrájuk az ősi emlősök meglévő struktúráinak módosulásával jött létre, pl. egy szárazföldi emlős - bálna átalakulás sem követeli meg az emlős testterv (body plan) drasztikus átalakulását.
Csak néhány evolúciós újítás (innováció) mint pl. a Chordaták gerinchúrja nem vezethető vissza már meglevő struktúrákra.
KÜLÖN FEJEZETET SZENTELÜNK MAJD NEKIK!
-
5. Testarányok drasztikus megváltozásai – allometrikus növekedés. Alapja a heterokrónia, egyes testrészek, szövetek fejlődésének időzítésének megváltozása és/vagy a sejtek érzékenységének megváltozása bizonyos növekedési faktorokra egy „őshöz” viszonyítva.
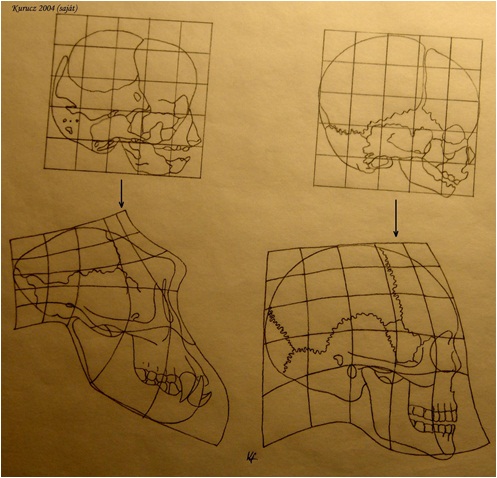
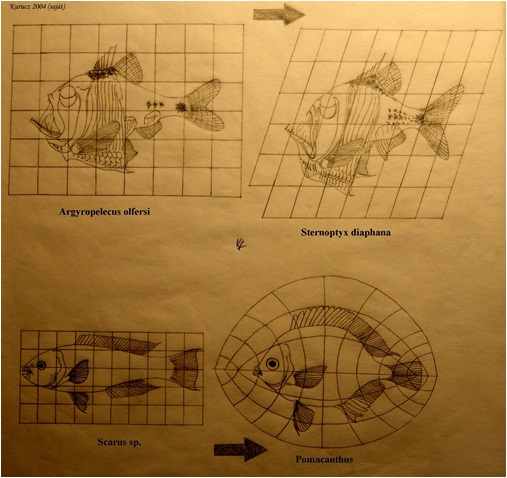
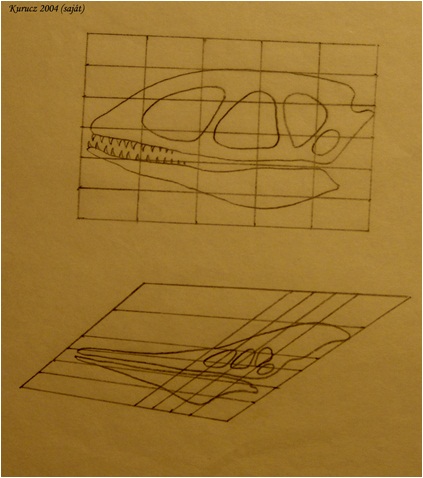
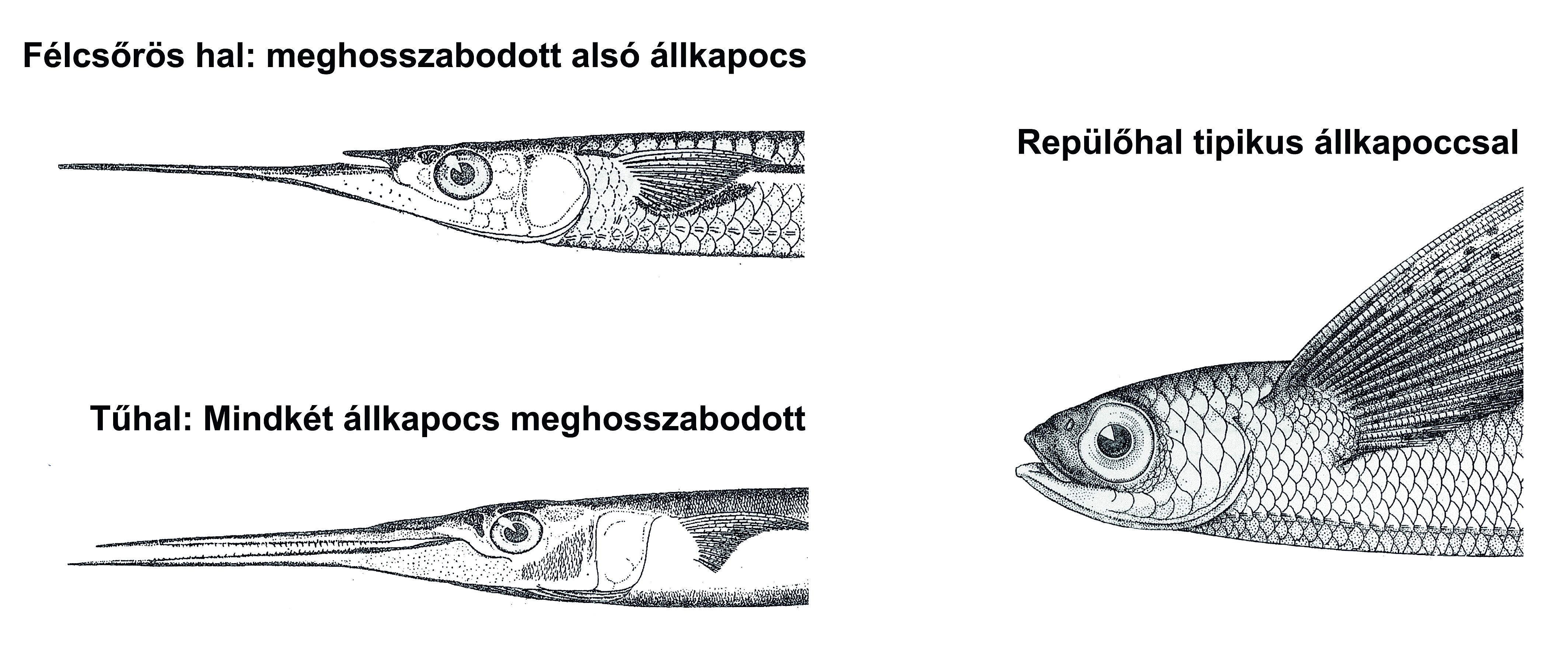
Általános elv, hogy a kis fejlődési változások nagy morfológiai változásokhoz vezethetnek. A növekedési ráta változásai drasztikusan változtathatják a formát - allometrikus növekedés jön létre. A változással statisztikailag-matematikai szempontból az allometria foglalkozik. Az allometria, vagy allometrikus növekedés az élőlény különböző részeinek, vagy dimenzióinak differenciális növekedését jelenti egyedfejlődése folyamán. (24. ábra; 25. ábra; 26. ábra) Az ember növekedésénél például a fej alacsonyabb rátával növekszik, mint a teljes test, míg a lábak magasabb rátával nőnek (23. ábra) Az egyes jellegek allometrikus rátájában történő változások – amiket „lokális heterokróniákként” értelmezhetünk -, igen fontos szerepet játszhatott az evolúcióban. Sok evolúciós változás pl. leírható azzal a megközelítéssel, hogy a helyi heterokróniák megváltoztatták egy vagy több jelleg alakját, pl. az ujjak megemelkedett megnyúlási rátájának figyelembevételével pl. levezethető a denevérszárny alakja, míg az alsó állkapocs alakja magyarázza a félcsőrös csuka, vagy a tűhal alakját (27. ábra)
4.22. ábra - Emberszabású majom és ember embrionális és felnőttkori koponyáinak allometrikus megfeleltetései.
 |

Az allometrikus növekedést gyakran írják le az y=bxa
Ahol x és y változók, mint pl. egy fog szélessége és magassága, vagy a fej és a test mérete. Az allometriás koefficiens, a leírja viszonylagos növekedési rátájukat. Ha a=1, a növekedés izometrikus, ami azt jelenti, hogy a két struktúra vagy dimenzió ugyanolyan rátával nő. Ha y gyorsabban nő, mint x, mint a testsúly és lábhossz viszonylatában a ›1, (pozitív allometria), ha y viszonylag lassan nő a‹1 (negatív allometria), mint a testhossz és fej esetében embernél. A később tárgyalandó pedomorfózis és a peramorfózis egyaránt lehet a fejlődés nagyobb rátájának, vagy hosszabb idejének az eredménye.
Az ír óriásszarvas (Megaceros giganteus) gigantikus agancsa, amely a testsúlyhoz viszonyítva a legnagyobb ilyen képlet, peramorfikus jelleg, ami az állat hosszasabb fejlődésével áll kapcsolatban.
Az ok, amiért az evolúció már létező „terveken” át zajlik, éppen az, hogy kis fejlődési változások nagy alaki változáshoz vezethetnek.
D’Arcy Thompson 1917-ben On Growth and Forma c. könyvében mutatta meg először, hogy drámaian különböző élőlények jönnek létre egyszerűen azáltal, hogy a test bizonyos részeinek növekedési rátája megnő.
-
Ez a fajta változás talán könnyen kifejlődik: a növekedés genetikai szabályozása viszonylag egyszerű – és a teljes testméret és a részek megfelelő módon alkalmazkodnak a későbbiek során.
-
Figyelembe véve, hogy modulárisan szervezett állatok alkotják az állatvilág legnagyobb fajszámú és legváltozatosabb morfológiájú csoportjait, talán van összefüggés a testszerveződés és az evolúciós diverzitás között. Egy lehetséges magyarázat az összefüggésre, hogy a moduláris szerveződés lehetővé teszi, hogy egy rész változzon anélkül, hogy más részeket befolyásolna. Azoknak a genetikai mechanizmusoknak az evolúciója, amelyek a részek individualizációját szabályozzák, lehetővé teszi, hogy a részek fejlődése „szétkapcsolódik”. Pl. a madarak mellső és hátsó végtagja egészen más módon fejlődik, és további módosulásuk is lehetséges.
A fejlődés változásain át zajló makroevolúció másik alapfeltétele a molekuláris parszimónia, az a jelenség, hogy bár a fejlődés nagyon különböző az élővilág különböző képviselőinél, a fejlődéshez mégis ugyanazokat a molekulákat használja minden vonal, azaz az élet takarékos, létezik egy „kis toolkit”.
A toolkit evolúciójának külön fejezetet szentelünk.
DNS és diverzitás
Egy élőlény minden sejtjében ugyanaz a genom van, hogyan fejeződnek ki más-más gének a különböző sejtekben?
Genomi ekvivalencia és epigenetika
A többsejtű élőlények egyedfejlődése során a megtermékenyített petesejtből, a zigótából számos, különböző működésű és szerkezetű sejtféleség alakul ki. Könnyen átláthatjuk a felmerülő kérdést és annak bonyolult szépségét: hogyan alakulhat ki a szervezet sejtjeinek sokfélesége egyetlen diploid sejtből: egyetlen sejtmagból, és az azt körülvevő citoplazmából? Milyen módon használ fel az embrió egyetlen genomot kiinduláskén t ilyen fokú változatossághoz?
Ha a morfológiai diverzitás a fejlődésről szól, a fejlődés pedig a genetikai szabályozó programok eredménye, a diverzitás evolúciója közvetlenül összefügg a genetikai szabályozó programok evolúciójával? Tulajdonképpen igen. A fejlődésbiológia azt tanulmányozza, hogy a megtermékenyített petesejt potenciálja hogyan változik a fejlődés során. Alapvető tétele az, hogy a sejtek differenciálódása a genetikai anyag mennyiségének és az azt felépítő bázisok sorrendjének megváltozása nélkül következik be. A testi sejtek – nagyon kevés kivételtől eltekintve – ugyanazt a génkészletet tartalmazzák, és a különféle sejtek egyazon génkészlet különböző génjeinek kombinációit hordozzák. A differenciális génexpressziós elmélet számos alátámasztást nyert, és egyaránt sokat köszönhet az embriológiának és a genetikának.
A differenciális génexpressziós elmélet kiterjesztése az epigenetika, amely a fejlődés minden vonatkozását figyelembe veszi. A fejlődés új, a sejtet alapvető egységnek, és az abban lejátszódó génexpressziós folyamatokat a fejlődés alapjának tekintő elméletét epigenetikának nevezzük. Az epigenetikai kontroll azoknak a genetikai faktoroknak az összessége, amelyek a sejtre hatva szelektíven szabályozzák a génexpressziót és a fejlődés folyamán egyre nagyobb fenotípusos komplexitáshoz vezetnek.
Bonyolult kérdések, hogy mit jelentenek a homológ szervek, ortológ és paralóg gének. Mára közhely, hogy az evolúciós változások visszavezethetők a genetikai anyag változásaira. Az evolúciós változásokat mindenképpen tükrözi az élőlény genetikai anyaga (genomja) és így a kódolt fehérjék spektruma. Az evolúciót szokás az allélgyakoriságok változásaként meghatározni a populációkban.
A homeobox nevű DNS-motívumot tartalmazó gének felfedezése minden állatcsoportban – egészen a Coelenterákig – minden eddiginél erősebb érv a genetikai folyamatosság és közös molekuláris szintű fejlődési elvek mellett. Az anatómiai struktúrák és fejlődési útvonalak diverzitása közös alapokból sarjad ki.
A morfológusok homológiákról beszélnek a közös evolúciós eredetű jellegek kapcsán. Pl. a különféle élőlények homológ szervei egy ősi szervből fejlődtek ki, ha eltéréseik miatt ez néha nehezen is felismerhető. A homológia koncepció megkülönböztet általános homológiát, azaz megfelelést különböző fajok szervei között és sorozat homológiát (serial homology) egy adott élőlény struktúrái (pl. mellső és hátsó végtag) vagy ismétlődő struktúrái (pl. szomiták) között. (A homológia fogalma és értelmezései a leginkább forrongó problémák között vannak a biológiában. Semmiképpen nem célunk a fogalom teljes letisztázása.)
Hasonló módon, a molekuláris biológusok, ha homológ génekről beszélnek nem-véletlenszerű (nem random) DNS szekvencia azonosságot értenek rajta a különféle élőlények génjei között. Nyilvánvaló, hogy a szekvencia azonosság magas foka közös leszármazás eredménye, pl. egy géncsalád tagjai biztosan egy ősgénből származtathatók le. Párhuzamosan a morfológusok terminológiájával a molekuláris biológusok beszélnek homológ vagy ortológ génekről, ha a nagyon hasonló szekvenciájú gének különféle fajokban, míg paralóg génekről, ha a hasonló gének egy élőlényben vannak jelen, mivel génduplikáció történt valamelyik ősben. Jelentős kérdés, hogy a genetikai anyag szintjén megjelenő homológiák megfelelnek-e a fenotípus szintjén megjelenő homológiáknak és fordítva: reprezentálnak-e a fenotípus homológiái örökítő anyag szintű homológiákat? A válasz sajnos nem. A homológ génekről átíródó fehérje molekulák a legkülönbözőbb szövetekben bukkanhatnak fel, olyanokban is, amelyek a morfológusok szerint nyilvánvalóan nem homológok. Pl. a Hoxd13 fehérje előfordul a kinövő farok, a láb és a mellső végtag csúcsán. A homológ fehérjék és homológ szervek között tehát nincsen kölcsönös megfelelés. Függetlenül a homológia nem egyértelmű használatától, vannak gének, amelyeknek meghatározó szerepet játszanak a sejttípusok, szövetek, szervek pozíciófüggő kidolgozásában. Az ilyen gének rendszerint transzkripciós faktorokat kódolnak, amelyek az alárendelt gének egész sorát irányítják. Az ilyen géneket szelektor vagy mestergéneknek nevezzük, és már sokszor szóltunk róluk. Az homológ/ortológ és paralóg mestergének, amelyeket különféle élőlényekben találtak, szintén a fejlődés szabályozásának azonos alapelvei mellett szólnak, amelyeket hangsúlyozunk a későbbiekben.
A rovarok és az emlősök szeme hagyományosan nem homológ, hanem analóg, azaz párhuzamos fejlődésű, de azonos funkciójú szervként él a köztudatban. Újabb eredmény, hogy mindkét szerv fejlődésének kezdeténél homológ gének állnak, így a „klasszikus” analóg szerv nézet legalábbis átgondolandó (Lásd a toolkitről szóló fejezetet is).
Új funkciók eredete – molekuláris szint
A pontmutációk és kromoszómamutációk általában csak megváltoztathatnak egy funkciót vagy átváltoztathatnak kissé eltérő funkcióvá. Teljesen új funkciók hozzáadása a génrepertoár expanzióját igényli duplikáció és poliploidia által, amit a duplikált gének divergenciája követ a szokásos mutációs folyamattal. A genom poliploidiával történő kiterjedése - legalábbis növényeknél – gyakori, amit az is mutat, hogy a páros kromoszómaszámok gyakoribbak a páratlanoknál (a gyakori poliploidia miatt).
Ha az össz-DNS mennyiség megnő, pár bázis szubsztitúció elég lehet egy génnél az új funkcióhoz. Hall – E. coliban egy új génfunkciót hozott létre.
A lac Z gén a ß-galaktozidáz funkciót kódolja, az ebg gén – egy másik ß-gal gén, nem fermentál laktózt, de a gént indukálja a laktóz.
A második enzim funkciója nem ismert. Hall – az ebg által kódolt enzimet megváltoztatta úgy, hogy laktobionátot (egy másik szubsztrátot) képes fermentálni. Egy másik mutáció során úgy változott meg a regulátor elem, hogy a gént konstitutív állapotba hozta (azaz állandóan kifejeződik). A végleges új funkcióhoz tehát 3 egymásra következő strukturgén-mutáció vezetett.
Az evolúció megállna a variáció kimerülése miatt, ha új variáció nem adódna a populációkhoz mutáció, rekombináció és migráció által. Végeredményben minden új variáció a gén- és kromoszómamutációktól származik.
Az élőlények életciklusa nagyon különböző lehet és a fejlődési események időzítésének különbségei az élőlény minden sajátságára kihatnak.A relatív növekedési ráták változásának hatásáról volt szó (allometrikus növekedés). Az életciklusok látványos megváltozása is fontos terület.
A legtöbb állatfaj lárvális fázison keresztül fejlődik. A lárvaállapot gyakorta nagyon specializált. Bizonyos fajok életciklusa megváltozott, lárvaállapotuk ideje vagy aránytalanul megnyúlt vagy megrövidült. Azt a jelenséget, hogy bizonyos állatoknál őseikhez viszonyítva megváltozik egyes jellegek megjelenésének ideje ill. fejlődési üteme, heterokróniának nevezzük.

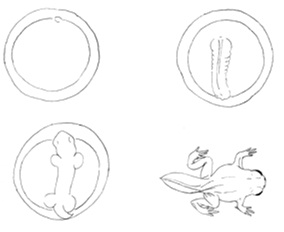
Neoténián a juvenilis formák megtartását értjük, ami abból ered, hogy a test fejlődése elmarad a gonádokéhoz viszonyítva, amelyek tartják az ősökre jellemző ütemet. Gould szerint a neoténia annak a módja, hogy a faj megszabaduljon a specializált felnőtt formától és képlékenyebb fiatal formák útján alkalmazkodjon más evolúciós irányokhoz (28. ábra)
A progenezis szintén a juvenilis forma megtartását fedi, de a gonádok és a csíravonalsejtek a normálisnál gyorsabban fejlődnek és szexuálisan éretté válnak, amikor az egyed teste még juvenilis.
A direkt vagy közvetlen fejlődéskor az embriókból a lárva állapot teljes kihagyásával közvetlenül kis felnőtt egyedek képződnek.
A mexikói axolotl (Ambystoma mexicanum) neoténiásan fejlődik, azaz nem megy át metamorfózison, mivel a hipofízise nem termel aktív tiroid stimuláló hormont (TSH). Ha az állatokba vagy tiroid hormont vagy TSH-t juttatnak, felnőtt egyed alakul ki. Az állat ilyen átalakulása a természetből nem ismert (29. ábra)
Az Ambystoma tigrinum egyes populációi közül a hidegebb vízben élők neoteniásak, míg a melegebb vidéken élők átalakulnak. Az előbbiek átalakulása elősegíthető melegebb körülmények között. A faj hipotalamusza úgy tűnik, hogy csak magasabb hőmérsékleten képes TSH-releasing factort termelni.
4.28. ábra - Az axolotl indukált metamorfózisa. A baloldali egyed az axolotl normális egyede, a jobboldalit tiroxinnal kezelték, előidézve metamorfózisát
 |
Más fajok neoténiásak maradnak hormonális beavatkozás után is. A Necturus és Siren fajok ilyenek. A Necturusból bizonyítottan hiányzik a TRb mRNS, a tiroxin-receptor RNS-e.
A neoténia mellett szól, hogy más fajok felnőtt egyedeivel azonos méretű lárva fejlődik ki, amely más fajokkal azonos korban is szaporodik.
Progenezis
A progenezis tette lehetővé bizonyos fajok számára új ökológiai fülkék (niche) elfoglalását. A Bolitoglossa occidentalis trópusi szalamandra, amely fákon él. Lábujjai között hártyák feszülnek, teste kicsi. A kis testet a lábak hártyái által kifejtett tapadási erő képes megtartani. A nyolcvanas évek elején kimutatták, hogy a Bolitoglossa occidentalis nagyon hasonlít két rokon faj fiatal egyedeire, amelyeknek termete kisebb, és ujjaik még nem nőtték túl a hártyákat. Úgy gondolják, hogy a B. occidentalis sokkal korábban vált érett felnőtt egyeddé, mint elődei. Sajátságai így alkalmassá tették a fán lakásra (30. ábra)

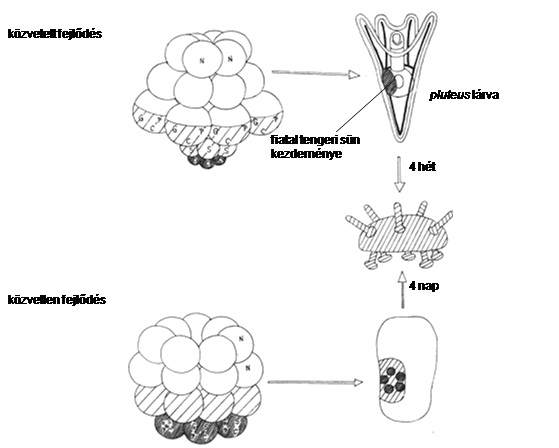
Közvetlen fejlődés békáknál
A közvetlen fejlődés szép példáit találhatjuk a lárvaállapotot „kihagyó” békáknál és a pluteus lárvát nélkülöző tengeri sünöknél.
Az Elentherodactylus coqui Puerto Ricon él. Az eddig említett békákkal szemben petéi a nőstény testén belül termékenyülnek meg. 31. ábra A peték átmérője 3,5 mm, térfogata kb. húszszorosa a Xenopuséinak. A peték lerakását követően a hím ráül a fejlődő embriókra, megóvva őket a ragadozóktól és a kiszáradástól. Korai fejlődésük nagyon hasonlít más békákéra, de nem sokkal a velőcső záródása után végtagbimbók jelennek meg, amelyekből végtagok alakulnak ki, tehát az állat „kihagyja” a lábatlan ebihal állapotot és kis farokkal rendelkező békává fejlődik. A végtagbimbók kialakulásához nem szükséges tiroxin. A béka fejlődéséhez tehát nem szükséges nagyobb mennyiségű víz, így annak hiányában is tud terjedni.
Közvetlen fejlődés tengeri sünöknél
A tengeri sünöknél az elsődleges mezenchima-sejtek betüremkednek és kiválasztják a pluteus lárva kalcium karbonátból álló vázát. A lárva addig nő, amíg a mikromerekből származó cöloma vezikulák kialakulnak a bél mellett. A bal oldali cöloma tovább növekszik és hidrocőlt alakít ki, ami a fölötte elhelyezkedő ektoderma invaginációját indukálja és kialakul a vesztibulum. A hidrocől és a vesztibulum együttesen kezdeményt formál, ami növekszik a lárvában mindaddig, amíg az ki nem bújik a metamorfózis folyamán és nem alakul fiatal tengeri sünné.
Bizonyos tengeri sün fajok fejlődéséből eltűnt a pluteus lárva. A közvetlen fejlődés és a szikben gazdag pete mérete között összefüggés állapítható meg: a 60-200 μm átmérőjű petével rendelkező fajok pluteus lárván keresztül fejlődnek. A 300-350 μm átmérőjű petével rendelkező fajoknál kialakul a pluteus, de annak belei nincsenek, így nem táplálkozik, fejlődése sokkal gyorsabb, mint az előző fajoké. Vannak fajok, amelyeknek petéje 2 mm átmérőjű, ezeknek a fejlődése közvetlen.
A jelenség érdekessége, hogy mind a békák, mind a tengeri sünök konzervatív lárvaállapottal bírnak, a konzervativitással járó kötöttségek feloldhatók úgy, hogy a petében lévő nagy mennyiségű szikanyag feleslegessé teszi a táplálkozó lárvát.
Molekuláris szint: bizonyos gének heterokróniája. Pl. tengeri sünöknél a közvetlenül fejlődő fajok „felnőtt” génjei sokszor korábban fejeződnek ki, lárvális génjei szupresszálódnak a közvetett fejlődésűekhez képest. Egy direkt fejlődésű faj wnt5 génje ugyanott kapcsol be, de korábban, mint közvetett fejlődésű rokonáé- a juvenilis struktúrák már az embrióban megjelennek a lárva helyett. (32. ábra)
A makroevolúciós változás mechanizmusai - barkácsolás a struktúrák és a génexpresszió szintjén
1977 – Jacob 33. ábra 1.) az evolúció azzal dolgozik, amije van – létező részeket kombinál újra, nem (vagy ritkán) „teremt” új részeket. Ő vezeti be a „tinkering” (barkácsolás) fogalmát az evolúció működésére vonatkoztatva.

Példa a strukturális szintű „tinkeringre”: az aortaívek változásai gerinces állatoknál
Az evolúció barkácsoló jellegére szép példát ad a különböző Chordaták keringési rendszereinek összehasonlítása, ahol nyomon követhető egy alapforma módosulása. 34. ábra
Köszönjük az alfejezetet Dr. Csoknya Máriának.
Szív: a rendszer központja.
Ütő- vagy verőerek (artériák): a szívből a kapilláris rendszer felé vért szállító erek rendszere.
Gyűjtőerek (vénák): a kapilláris rendszer felől a szívbe vért szállító erek rendszere.
Hajszálerek (kapillárisok): az artériás és a vénás rendszert összekötő legkisebb átmérőjű erek.
AZ AORTAÍVEK
A gerinces állatok artériás rendszerét és változásait a lándzsahal artériás keringési rendszeréből vezethetjük le. Mint azt az összehasonlító anatómiai kutatások eredményei mutatják, a lándzsahal keringési rendszere kiváló alapanyagnak bizonyult az evolúciós barkácsolás számára.
Lándzsahal vérkeringése és aortaívei (33. ábra) A gerincesek zárt vérkeringésének fejlődése leginkább az egyre tökéletesedő szív evolúciójával korrelál. Az anatómiai tinkering a “szívtelen” lándzsahaltól indul és vezet a csontoshalak két-, a kétéltűek három- és a madarak/emlősök négyüregű szívéig. Az anatómiai szintű barkácsolás eredményeként az oxigenálódott vér egyre kevesbé keveredik a testből a vénák által szállított CO2-dal telített vérrel. A négyüregű szívben az artériás és vénás vér tökéletesen szeparálódik, ami az állandó testhőmérséklet fenntartásának lehetőségéhez vezet és további evolúciós lépések előfeltételének tekinthető.
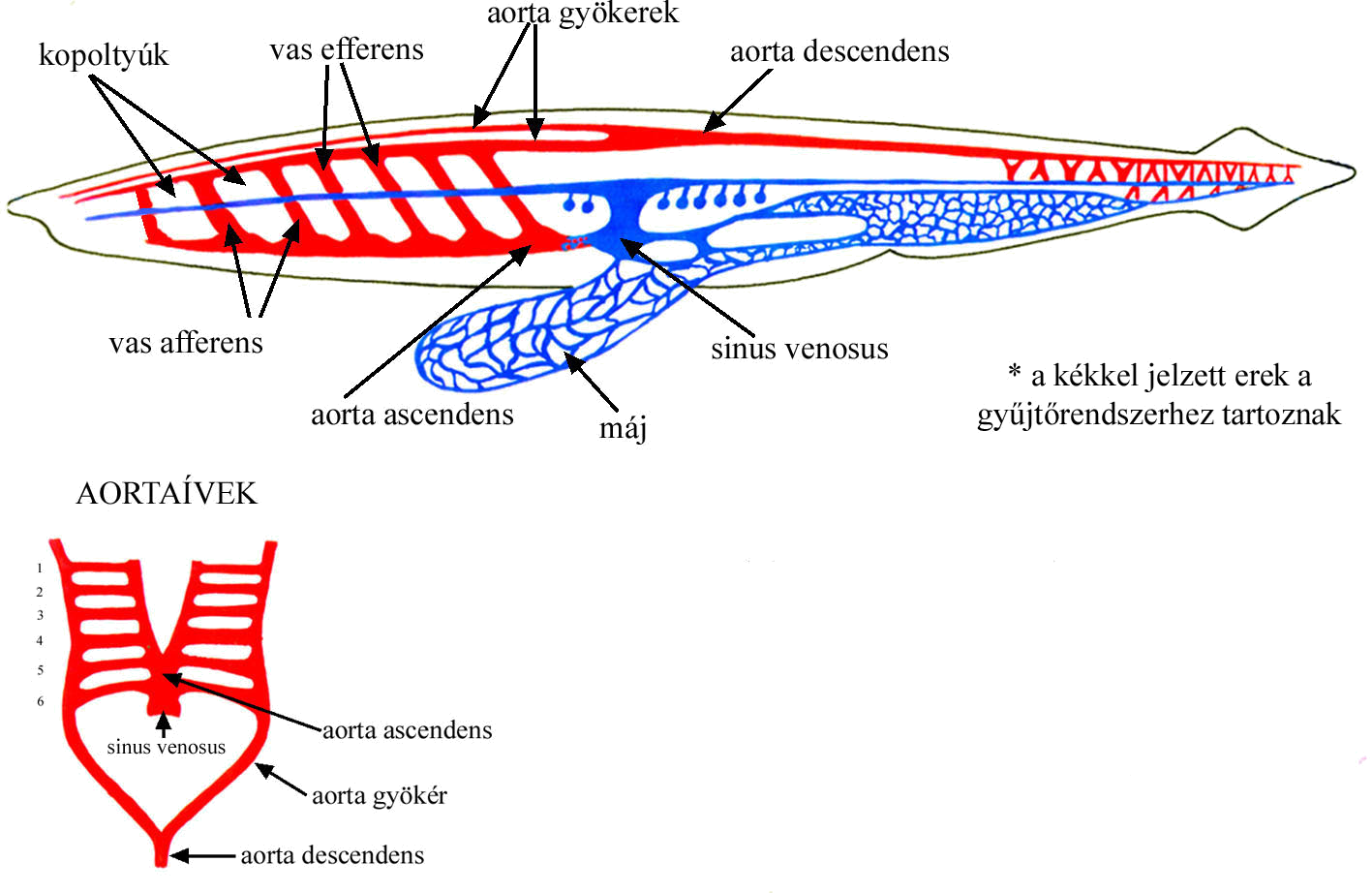
Keringésének jellemzői:
1. Nincs igazi szíve.
2. Hat pár kopoltyúja, és így 6 pár aortaíve van.
3. Egy vas afferens és egy vas efferens egy kopoltyúív artériát ad.
4. A kopoltyúív artéria megnevezés helyett, röviden csak az aortaív kifejezést használjuk.
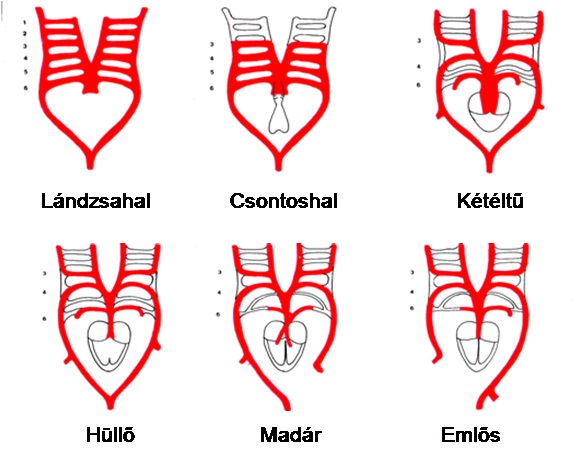
ÖSSZEFOGLALÓ (34. ábra)
-
A lándzsahal 1. és 2. kopoltyúív artériája minden gerinces osztály képviselőinél obliterálódik.
-
Halaknál a 3 - 6. kopoltyúív artériák továbbra is megmaradnak.
-
A lándzsahal 3. kopoltyúív artériájából a kétéltűektől az emlősökig fejütőerek lesznek.
-
A lándzsahal 4. kopoltyúív artériája kétéltűeknél és hüllőknél szimmetrikus aortaív lesz. Madaraknál az ív jobb, emlősöknél az ív bal oldali része marad meg aortaívnek; ebből ágazódnak ki a megfelelő oldali végtag artériák. Az aortaív másik oldala a. subclavia lesz.
-
A lándzsahal 5. kopoltyúív artériája kétéltűektől obliterálódik.
-
A lándzsahal 6. kopoltyúív artériája kétéltűektől minden osztályban szimmetrikus tüdőartériaként funkcionál.
1940 - Richard Goldschmidt - kis genetikai változások felgyűlése nem elégséges az evolúciósan új struktúrák (dúcléc, fogak, teknőshéj, tollak, stb.) létrejöttéhez - az ilyen változásokhoz a fejlődést szabályozó gének öröklődő megváltozására van szükség!
Évtizedekig az ilyen génekről csakis a hipotézisek szintjén gondolkodtak. Jacob szerint a „barkácsolás” az embriók létrehozásában fontos géneknél a legvalószínűbb , nem a felnőtt egyedben funkcionáló géneknél.
Wallace Arthur (2004) összegyűjtötte azokat a módozatokat, ahogy Jacob „tinkering”-je létrejöhet a génexpresszió szintjén:
- Heterotópia (változás az adott gén kifejeződésének lokalizációjában)
- Heterokrónia (változás az időzítésben)
- Heterometria (változás a mennyiségben)
- Heterotipia (változás a minőségben)
Mindezen változások előfeltétele, hogy a génkifejeződési mintázatok MODULÁRISAK legyenek, azaz a mintázat különböző elemeit különböző enhenszer elemek szabályozzák- egyes elemek változása így nem feltétlenül befolyásolja egy másik elem fejlődését
A fejezet és az anyag további részében jónéhány példát láthatunk a jelenségre, a gének kifejeződésének lokalizációs változásaira. Itt csak néhány látványos példát sorolunk fel
A kacsa úszóhártyáinak kialakulását a gremlin gén kifejeződése tette lehetővé az interdigitális térben. A Gremlin fehérje a BMP-k (bone morphogenic factor) gátlója. A BMP-ket gondolják felelősnek az apoptózis beindításáért – hiányukban nincsen sejtelhalás
Denevéreknél hasonló a helyzet, csak náluk az FGF8 fejeződik ki az interdigitális térben a BMP-k gátlójaként
Az emlősök fogai
Stephen J. Gould (1989): a paleontológusok szerint az emlősök evolúciójának lényege, hogy két fog párosodik, és létrehoz egy kicsit megváltozott fogat. Az zománc (enamel) marad meg a legtovább, tovább mint a csontok, és a változása kétségtelenül fontos tényező az emlősök sikerében.
Jernvall és Salazar- Ciudad (2000, 2002) – Geographical Information System (GIS)- génkifejeződés elemzésére a zápfogkezdeményekben – kimutatták, hogy a génexpressziós mintázatok előrevetítik a foggumók helyét - sőt a különbségeket a pockok és egerek fogmintázata között.
2004: matematikai reakció-diffúziós modellt javasolnak 35. ábra:
4.35. ábra - A fogak alakjának megváltozása a fosszíliákban és egy elfogadott fejlődésbiológiai modell (a reakció-diffúziós modell) alapján
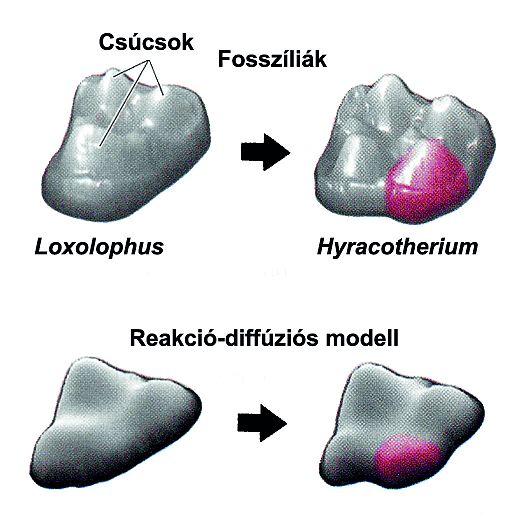 |
A Shh (sonic hedgehog) és FGF (fibroblast growth factor; a zománckezdemények jelközpontjai termelik) gátolják a BMP termelődését, míg a BMP stimulálja saját és inhibitorai termelődését. Így aktivátor régiók alakulnak ki (BMP-hatásra), amelyek gátolják az epitélium sejtjeinek szaporodását és inhibitor régiók (FGF, Shh-hatásra), amelyek blokkolják a BMP szintézisét, és ettől függetlenül stimulálják a mezenchimális proliferációt.
Eredménye egy olyan dinamikus génexpressziós mintázat, amely változik, ahogy a fog alakja változik, ill. egy fogforma, amely úgy változik, ahogy a génexpressziós mintázat.
A modell megengedi a paraméterek változását a fog fejlődése során. A diffúziós ráta és a kötési konstans apró változásai komoly morfológiai változásokhoz vezetnek.A modell megjósolja bizonyos fogtípusok gyakori kialakulását és megerősíti a paleontológusok konklúzióit.
Egyéb példák: a rákok morfológiai változatosságát a homeotikus gének kifejeződésének különbségei okozzák (ld. előbb). A madártollak kialakulásában biztosan szerepe volt a Bmp2 és az Shh gének kifejeződési doménjainak változásainak (ld. később). A gerincesek gerinccsigolyáinak alakulása a Hox-gének kifejeződésének változásával mutat korrelációt (ld. később is)
A jelenséget több helyen említettük és több-kevesebb részletességgel tárgyaltuk. A heterokrónia a fejlődési folyamatok időzítésének megváltozása a generációk folyamán. Kissé leegyszerűsítve egyik modul megváltoztatja kifejeződésének idejét a többi modulhoz viszonyítva.
Szép példa a madarak és gyíkok szeme - korábban kezdik meg a fejlődésüket, mint az emlősök, így filotipikus stádiumú embriójuk (pharyngulájuk) szeme nagyobb (36. ábra).
Erszényesek szája és mellső lába gyorsabban fejlődik 37. ábra
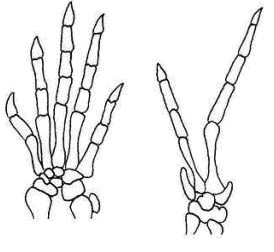
Hemiergis gyík genusban vannak 3, 4, 5 ujjú fajok- attól függően, hogy meddig aktív shh (sonic hedgehog) gén a végtagbimbó ZPA régiójában (38. ábra)
Heterokrónia és az állkapcsok eredete
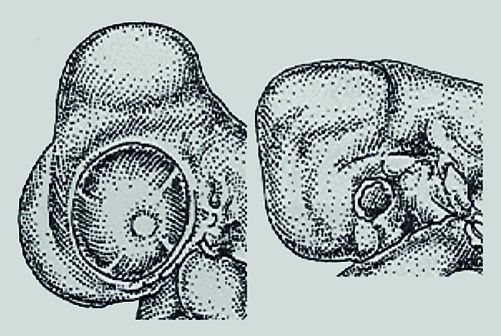
Heterokrónikus változások felelősek az egyik legfontosabb gerinces adaptáció, az állkapocs kialakulásáért. Amikor a gerincesek feje először kialakult, hasonlítottak a Cyclostomatákhoz: teljes vázuk porcos volt, fejük hasonlított egy csigolyához. A Gnathostomatáknál (állkapcsosok) a kraniális dúclécsejtek belépnek a zsigeri ívekbe, és amelyek az első ívbe lépnek, alakítják ki a mandibulát (alsó állkapocs) és a maxilláris nyúlványt (szájpad).
A Cyclostomatáknál a dúclécsejtek ugyan vándorolnak, de az I. pharyngeális ív nem alakítja ki a Meckel-porcot és a belőle származó mandibulát. A dúcléc-sejtek - ehelyett - egy kerek, állkapocs nélküli szájat alakítanak ki. Két feltételnek kellett teljesülnie, hogy az első zsigeri ív dúclécsejtjeiből állkapocs legyen:
(i) permisszív környezet (ii) új instrukciók
-
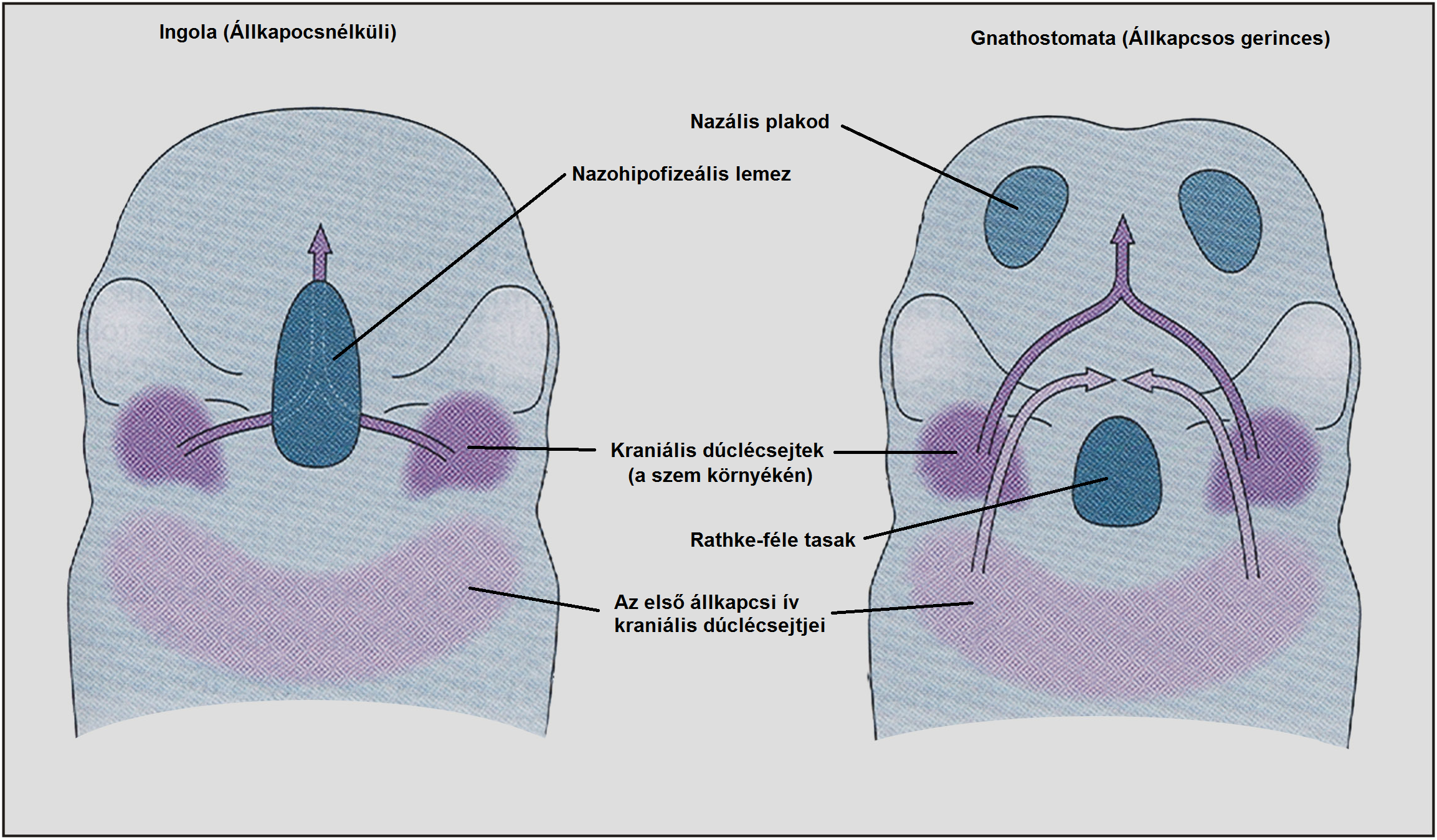
A permisszív környezet úgy alakult ki, hogy eltűnt egy gát (39. ábra). A Cyclostomaták naso-hypophiseális lemezéből alakul ki a nazális epitélium és az agyalapi mirigy. Ez az epiteliális lemez a szó szoros értelmében akadályt képez a dúclécsejtek számára - csakis alatta vándorolhatnak - a vándorló sejtek aztán kialakítják a Cyclostomaták ajkának felső ajkát.
A gnathostomatáknál a naso-hypophyseális lemez korán nazális plakodokra és Rathke-féle tasakra hasad - ezek között a képletek között már vándorolhatnak a dúclécsejtek. A két struktúra között rostrálisan vándorló dúcléc-sejtekből kialakuló struktúra a mandibula
4.39. ábra - A nazohipofizeális lemez felhasadása új lehetőséget teremt a sejtvándorlásokhoz, így új képletek alakulnak ki
 |
(ii) Új instrukciók: Hox-gének kifejeződése is elcsúszik. Cyclostomatáknál az I. zsigeri ívben Hox-gének kifejeződnek, a Gnathostomatáknál nem. Ha a Hox-géneket kifejeztetik, az állkapocs kialakulása gátlódik => gátlásukra szükség van az új instrukciók megjelenéséhez.
A heterometria egy struktúra, vagy géntermék mennyiségi változását jelenti
A Darwin-pintyek speciációjában fő lépése a kaktusz- és földipintyek elválása. A földipintyek csőre nagy és széles, magvakat törnek vele. A kaktuszpintyek csőre keskeny, hegyes: rovarokat és kis virágrészeket csippentenek ki vele a kaktuszvirágokból. A fajok közötti különbségeket a frontonazális nyúlvány dúcléc-eredetű mezenchimájának növekedési változásai okozzák. Mint Abzhanov és mtsi (2004) megállapították, korreláció áll fenn a csőr formája és a Bmp4 fehérje mennyisége között: a földipintyeknél koraibb és erőteljesebb a kifejeződés. Csirkék: Bmp4 kifejeződés növelése=> széles, erős csőr; BMP-inhibitor => keskeny, hegyes csőr.
Heterometria – IL4
Emberi IL4 - 524T allél humán vonalon jött létre, hajlamosít allergiára, asztmára, contact dermatitisre, stb. Az új allél egyel több NFAT (nuclear factor for activated T cells) transzkripciós faktor kötőhelyet tartalmaz, ami háromszoros IL4-fehérje mennyiséget eredményezett. Bélparaziták ellen kedvező lehet az új allél, szelekciót is mutattak ki bizonyos populációkban, amelyek ki vannak téve bélparazitáknak. Az allometria is a heterometria egyik formájaként (is) értelmezhető.
Allometria
A modularitás következménye az allometria: az élőlény részeinek különböző rátájú növekedése. Leggyakrabban a célsejtek növekedési hormonra való érzékenysége, vagy a termelt növekedési hormon mennyisége változik meg, de a növekedés időbeliségének megváltozásával is csaknem törvényszerűen kapcsolatban van.
Pl. a chondrociták közötti helyi különbségek okozták, hogy az ősi lovak középső ujja 1,4X gyorsabban nőtt, mint a többi ujj (ld. Bevezető fejezetben is)
Pl. 4-5 mm-es bálnaembriónál még az emlősökre jellemző helyen az orrlyuk (40. ábra) , vagy a gerinces végtagok kialakulása (41. ábra).
Az allometria úgy hozhat létre evolúciós újdonságokat, hogy átmegy bizonyos küszöbökön („bifurkációs pontokon”)
4.40. ábra - Az orrcsont és állcsont látványos allometriás növekedése az orrlyuk fejtetőre helyeződését eredményezi bálnáknál
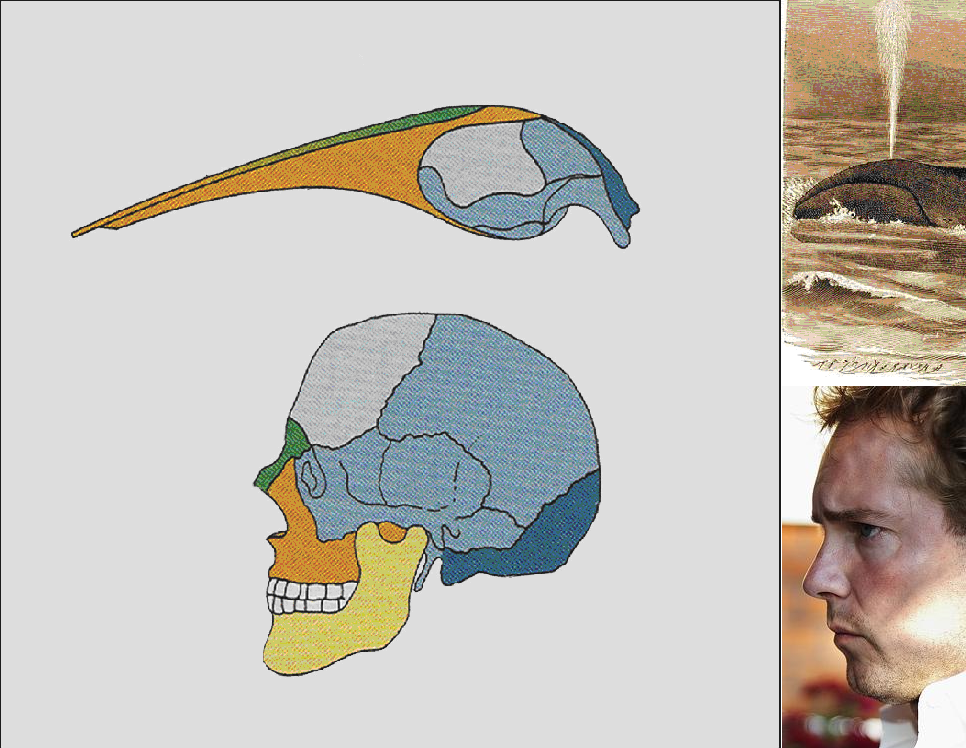 |
4.41. ábra - A röpképtelen új-zélandi kiwi és egy denevérfaj végtagjainak illetve azok részeinek allometrikus növekedése
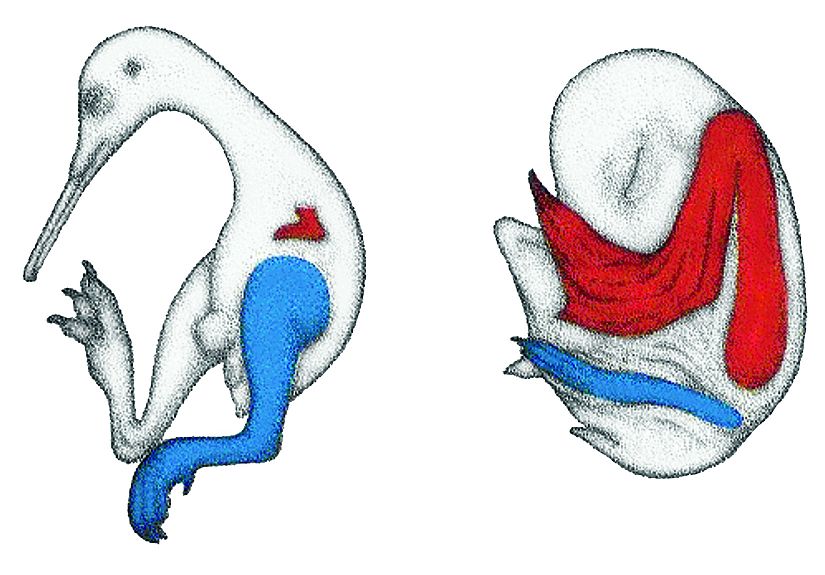 |
Heterokróniánál, heterotópiánál, heterometriánál a mutációk a gének regulátorszakaszát érintik - a tulajdonképpeni géntermék ugyanaz marad, akkor is, ha új helyen, új időben, vagy más mennyiségben szintetizálódik. A heterotipiás változás a gén fehérjekódoló szakaszát érinti, így a fehérje funkciói változnak meg. Az egyik látványos példa: a rovaroknak mindössze hat lába van, szemben a többi Arthropodával, amelyeknek rendszerint jóval több. A jelenség kulcsa az Ultrabithorax fehérje és a Distalless kölcsönhatása: míg a többi ízeltlábúnál az UBX nem gátolja a Dll-t, a rovaroknál az eredeti 3’ véghez egy 10 alanint kódoló szakasz került, az így kódolt fehérje pedig represszálja a Dll-t.
A heterotípia tehát jelentős tényező az „új” szabályozó fehérjék létrejöttében. Amellett, hogy természetesen létrejöhet „általános” mutációs mechanizmusokkal, a „replikációs megcsúszás” (a mechanizmusra inkább rávilágító név, mint láthattuk, a „slipped mispairing” (42. ábra)) jóval gyakoribb módja az új fehérjék létrejöttének. Leggyakoribb, de mindenképpen legnagyobb eséllyel hatásos módja, hogy egy kodon válik sokszorosan ismétlődővé, így a fehérjében egyazon aminósavból lesz egy hosszú sorozat, ami megváltoztatja a fehérje sajátságait. A Hox-család például emberben a leginkább poliadenin-gazdag fehérjecsoport. A glicin-ismétlődések gyakoriak a külső megtermékenyítésű tengeri fajok gaméták kapcsolódását elősegítő fehérjéiben. Az ismétlődések hossza az új pontmutációkkal keletkező allélokhoz viszonyítva100 000-szeres rátával keletkeznek.
Számos transzkripciós faktorban találtak glicin-, prolin-, vagy alanin-ismétlődéseket. Kutyákban meg lehetett állapítani összefüggést az Alx4-gén kis ismétlődés-számú alléljának jelenléte és bizonyos morfológiai jellegek között. A pireneusi juhászkutya az egyetlen fajta, amely homozigóta a kis ismétlődés-számú allélra, és az egyetlen, amelyre jellemző a polidaktília. A kivételes, normális ujjszámú egyedek más fajtákra jellemző allélokat hordoztak homozigóta formában. A glutamin és alanin tandem ismétlődéseinek különböző száma a Runx2 –génben korrelál a pofa sajátságaival (43. ábra). A tandem ismétlődésekkel sokkal jobban lehet magyarázni a kutyafajták evolúciós mértékkel gyors kialakulását az elmúlt, legfeljebb 135 000 évben (a pontmutációk rátája 10-7-10-9-re tehető).
A replikációs elcsúszásra mint „a természet kísérletezésére” („nature’s experimentation”) szoktak hivatkozni. Hatása sokszor végzetes, vagy káros: A Huntington-szindrómánál a CAG-ismétlődések száma szabadul el, és eredményez glutamin-sorozatokat, amelyek kölcsönhatásba lépnek a TBP(TATA-binding protein)-fehérjével, és akadályozzák az átírást. A törékeny-X szindróma is replikációs elcsúszásra vezethető vissza.
4.42. ábra - A rövid ismétlődő szakaszok esetén gyakran fellépő "szálelcsúszás"-ból eredő mutáció (slipped mispairing vagy strand slippage)
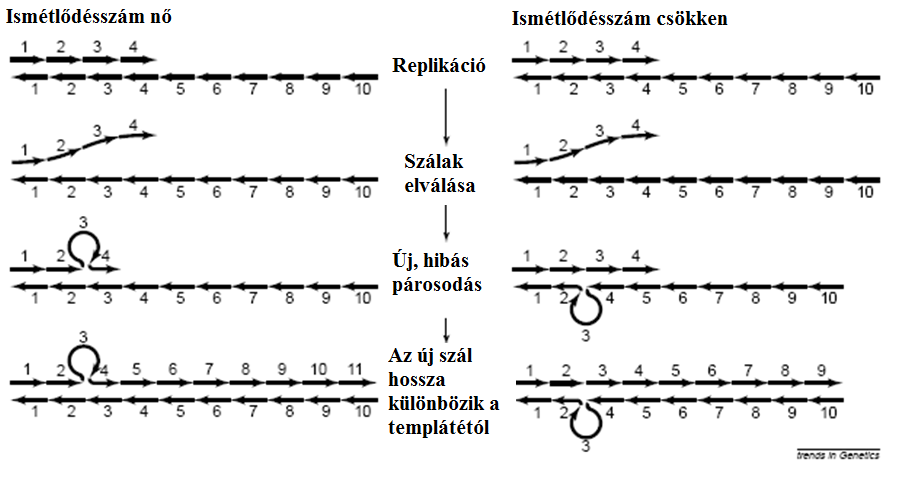 |

Toborzás
Az is előfordulhat, hogy már létező egységek, néha egész jelátviteli útvonalak toborozhatók új funkcióhoz. Szép példái a toborzásnak (recruitment):
1. Egy teljes, a szegmentációban használatos jelátviteli rendszer kooptálása a pillangó foltjának kialakulásához.
2. Az egérbe átvitt béka Distall-less gén (egészen pontosan szabályozó elemei + lacZ riportergén) kifejeződik mindenhol, ahol a békában + a bajuszszőrökben.
Az éjszaka aktív (nokturnális) állatok a rendelkezésre álló minimális fény maximális hasznositását néhány egyedülálló anatómiai es molekuláris szintű megoldással érik el. Az éjszakai látás tulajdonságának a megszerzése az anatómiai és gén/genom-szintű barkácsolás egyik legszemléletesebb példája. (44.ábra)
4.44. ábra - Kromatin-szintű barkácsolás a nokturnális állatok retinájában. A panel – Retina keresztmetszete. ↑ - A fény érkezési iránya. Narancssárga sejtek – Ganglion sejtek. Zöld sejtek – Interneuronok rétege. Piros sejtek – Pálcikasejtek. PS: Pálcikasejt sejtmagja, GS: Ganglionsejt sejtmagja. B panel – Pálcika- és ganglion-sejtek sematikus kromatin organizációja. C panel – Pálcikasejtek kromatin állományának átrendeződése az egér pálcikasejtjeiben, újszülött, egy hetes, két hetes, három és négy hetes egérben. Valamennyi panelre érvényes színkód: Zöld – eukromatikus, kék és piros – heterokromatikus régiók.
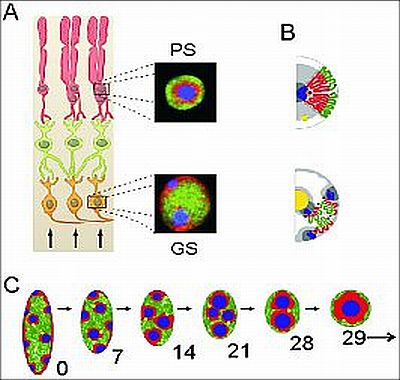 |
1. A nokturnális állatok retinájában az alacsonyabb intenzitású fény érzékelésére alkalmas fotoreceptorok, pálcikasejtek nagyobb arányban fordulnak elő, mint a nappal aktív (diurnális) állatok esetében.
2. Nokturnális állatok a retinája mögött egy fényvisszaverő réteg (tapetum lucidum) található, ami az első körben nem hasznositott fotonokat visszairanyitja a fotoreceptorok rétegebe.
3. A palcikasejtek egyedi sejtmagi szerveződest mutatnak, ami alapvetoen elter a test tobbi sejtjenek magjaban talalhato kromatin elrendezestol. Az altalanosan elterjedt kromatin topologia az interfazisos sejtekben úgy szerveződik, hogy a heterokromatikus regiok tobbnyire a sejtmag membranja menten, un. perinuklearis pozicioban vannak, mig a transzkripcionalisan aktiv, kevesbe zart eukromatikus regiok a sejtmagok kozepen lokalizalodik. A palcikasejtekben pontosan forditott topologiat mutat a kromatin, az eukromatin a periferian, mig a heterokromatin a sejtmag kozepen talalhato. Ennek a forditott (invertalt) kromatin elrendozodesnek optikai kovetkezmenyei vannak. A fény csak kis mertekben szorodik es fokuszalodik a fotoreceptor sejt azon reszere, ahol a fényerzekelo molekulak (rodopszin) tarolodnak. Tehat, egy olyan kromatin struktura alakult ki, ami optikai lencsekent funkcional, amellett, hogy maradektalanul ellatja az “egyeb” gen szabalyozassal kapcsolatos funkcioit. A retina fejlodesenek korai fazisaban a palcikasejtek magjai meg nem rendelkeznek ezzel a mikrolencse strukturaval es csak lepesrol lepesre alakul ki ez a specialis sejtmagi architektura. Egerek eseten a szuletest koveto 13. napon, a szemek kinyilasanak idopontjaban, mar javaban folyik az atrendezodes, ami az elso honap vegere fejezodik be. A fény szerepe a kromatin-alapu mikrolencse kialakitasaban, a tinkering iranyitasaban meg teljesen nyitott kerdes.