Tartalom
1894 fontos év az egyetemes biológia fejlődése szempontjából - Wilhelm Roux 1. ábra

deklarálta a KÍSÉRLETES EMBRIOLÓGIA létrejöttét. Tudta, az új megközelítéssel sok szálat elszakít az EVOLÚCIÓ és az EMBRIOLÓGIA között. Ígérete:„an ontogenetic and phylogenetic developmental mechanics to be perfected” („egy egyedfejlődési és törzsfejlődési mechanikára derül fény”), azaz a végeredmény az lesz, hogy a két terület ismét találkozik. Úgy gondolta, hogy az egyes embriók „fejlődésmechanikája” gyorsabban megismerhető, mint az evolúciós szint, de„ in consequence of the intimate connections between the two many of the conclusions drawn from the investigation of individual development throw light on the phylogenetic process” („a kettő közötti meghitt kapcsolat következtében sok olyan következtetés, amelyet az egyedi fejlődésből vonnak le, megvilágítja a filogenetikai folyamatot”). Roux próféciája jó száz év múlva teljesült:
A FEJLŐDÉSBIOLÓGIA visszatért az EVOLÚCIÓS BIOLÓGIÁHOZ, és kialakult az EVOLÚCIÓS FEJLŐDÉSBIOLÓGIA (= EVOLUTIONARY DEVELOPMENTAL BIOLOGY = EVO-DEVO), amely kulcs a földi diverzitás megmagyarázásához. Az EVO-DEVO integrálja a FEJLŐDÉS-, ÉS a POPULÁCIÓS GENETIKÁT- egyfajta „új szintézisként” értelmezhető.
Az új evolúciós szintézis alapelve:
- AZ EVOLÚCIÓ ALAPJA AZ ORGANIZMUS FEJLŐDÉSÉNEK ÖRÖKLŐDŐ VÁLTOZÁSA. Maga Darwin is hasonló nézeteket vallott, sőt, előtte is számosan
- A nézet összeegyeztethető a populációs genetikai felfogással: az evolúció oka eszerint a génfrekvenciák változása a populációban
Az EVO-DEVO a fejlődés és az evolúció kapcsolatát tanulmányozza. Célja azoknak a mechanizmusoknak a megértése, amelyeknek szerepük volt a fejlődés evolúciójában. A megközelítés természetesen két irányból történhet: a fejlődési folyamatok irányából (fejlődési folyamatok időzítése, új szöveti kölcsönhatások szerepe egy új morfológiai sajátság kialakulásában, ami jellemző lehet egy taxonra), és az evolúciós folyamatok irányából (pl. milyen szelekciós nyomás támogatja az új morfológia kialakulását).
Két kérdés vitán felül kiemelkedően fontos:
(1) Milyen szerepet játszott a fejlődés evolúciója a földi élet alakulásában?
(2) Hogyan befolyásolják a fenotípust befolyásoló fejlődési folyamatok a variációk létrejöttét és az evolúciós változást?
A természetes szelekció a fejlődés által létrehozott fenotípusokon hat, de a végső cél, hogy megértsük: miképpen befolyásolja az evolúciós potenciálokat és pályákat az a mód, ahogy a fejlődés létrehozza a fenotípusokat. A forma a fejlődés során jön létre. A morfológiai struktúrák végleges formája növekedéssel alakul ki.
„Evolúciós” változások egy szerv formájában “fejlődési” változások is egyben: a fejlődési folyamatok időzítésének és rátájának változása vezet hozzájuk. A szerv megnőhet, ha növekedése felgyorsul; alakot változtathat, ha egyes részeinek növekedési rátája felgyorsul - ilyenkor allometrikus növekedésről beszélünk.
Az evolúciós változások és az egyedfejlődés tehát elválaszthatatlanok.
A téma maga, a fejlődés és az evolúció kapcsolata régi, a már „klasszikus” elméletek közül is több, pl. a rekapituláció is ide tartozik.
A témakör, stílusos szóval élve, „transzformálódott” az utóbbi két évtizedben. A legutóbbi 20 év legfontosabb felfedezései közé tartozik, hogy a legtöbb állat - tkp. az összes, bizonyos komplexitás felett álló soksejtű, függetlenül attól, mennyire különböznek formailag – testszerveződésének (testtervének) fő vonatkozásait specifikus géncsaládokba tartozó, kisszámú gén szabályozza. Ennek az állati fejlődés „toolkit”-jeként emlegetett genetikai készletnek a felfedezése döntően befolyásolta a kutatást.
Döntően két kihatás emelendő ki:
1.) Lehetővé tette a biológusok számára, hogy felfedezzenek és felderítsenek széleskörűen konzervált molekuláris, celluláris és fejlődési folyamatokat, amelyeket korábban elfedtek az addig összehasonlíthatatlannak” vélt anatómiai viszonyok
2.) Az állati sokféleség tanulmányozását a „toolkit” gének
számának, regulációjának, funkciójának változására irányította az állati evolúció során.
A „toolkit gének tér-, és időbeli kifejeződési rendjének változásai az embriogenezis során összekapcsolhatóak a felnőttkori morfológia változásaival is.
A „TOOLKIT” FEJLŐDÉSRE SZAKOSODOTT GÉNJEINEK KIALAKULÁSA TALÁN MEGNYITOTTA AZ EVOLÚCIÓ ÚJ ÉS HAJLÉKONYABB MÓDJAIT: AZ ÉLET „EVOLVABILISEBB” LETT.
A fejlődés és a fejlődést szabályozó gének változásai a morfológiai evolúció alapjai. A morfológiai struktúrák a fejlődés során jönnek létre az egyes egyedekben. Ha az egyik faj másikká alakul, a fejlődési folyamat is megváltozik. A FEJLŐDÉS ÉS EJLŐDÉSGENETIKA „EVOLÚCIÓS” VÁLTOZÁSAI AZ ALAPJAI AZ EVOLÚCIÓS SZINTŰ MORFOLÓGIAI VÁLTOZÁSOKNAK. A fejlődés evolúciójának megértése nélkülözhetetlen a morfológiai evolúció megértéséhez (a molekuláris vagy kromoszomális szintű evolúció megértéséhez nem kellenek feltétlenül fejlődési ismeretek).
A biológusok már a XIX. században (elsősorban a franciák már a század legelején) megértették, hogy a fejlődés a kulcs az evolúcióhoz – a XIX. századi tudósoknak bizonyos eredményei, témafelvetései máig hatnak, máig „tankönyviek” (von Baer, Cuvier, Geoffroy Saint- Hillaire, Haeckel, Darwin, stb.)
Az utóbbi 15 évben tolódott a hangsúly a „toolkitre”: az EVO-DEVO talán fő témája: hogyan változott a „toolkit” a fajok között? A régi elméletek értéke vitathatatlan, de az újakkal való integrációjuk nem tökéletes- sokszor azok a gének, amelyek a régen tanulmányozott struktúrák, szervek, élőlények, fejlődési lépések fejlődését befolyásolják, még nem kerültek azonosításra.
Summa-summárum: A XIX. sz. elejétől napjainkig összegyűlt ismeretek hozzájárulnak a morfológiai evolúció megértéséhez. A szintézis fő forrásai: - Paleontológia- összehasonlító anatómia, embriológia - filogenezis kutatása (szisztematika, rendszertan)- genetika, molekuláris biológia, FEJLŐDÉSGENETIKA, FEJLŐDÉSBIOLÓGIA -EVOLÚCIÓ általában
Evolúciós perspektíva nélkül a fejlődés mikéntje legtöbbször értékelhetetlen!
A gerinceseknél pl. a korai fejlődés nagyon különböző, de beletorkollik egy sajátos, a legkülönbözőbb gerinces fajoknál szinte megkülönböztethetetlen stádiumba - majd divergálódik! Ez a különös filotipikus stádium, amely a neuruláció és szomitakialakulás után alakul ki, valamely távoli gerinces ősre jellemző!Megmaradt, sőt alapvető sajátsága a gerinces fejlődésnek - de az előtte és utána lévő szakaszok különbözőképpen zajlanak. Az ilyen változások a fejlődést irányító gének változásai miatt következnek be - új a környezethez jobban alkalmazkodó és sikeresebben szaporodó formákat az evolúció szelektálja ki.
A fejlődésbiológia egyik alaptémája a fejlődési mechanizmusok konzerválódása távoli rokon fajok között pl. Hox-gének, jelátviteli útvonalak a szélsőségekig konzerváltak, amiből az következik, hogy ha egyszer egy mechanizmus kialakult - újra és újra felhasználódik, ami az [evolúciós (fejlődés)] biológia sava-borsa. (Ezeket a témákat a későbbiekben részletesen kifejtjük.)
A következőkben a „klasszikus” eredményeket és elméleteket ismertetjük, amelyek megtermékenyítőleg hatottak a modern kutatók gondolkodására is, sőt, mit sem veszítettek aktualitásukból.
Máig ható eszmék: „Unity of type”, „conditions of existence”, és von Baer csodálatos megfigyelései
A XVIII- XIX.században két alapvetően eltérő nézetkör alakult ki:
1.) Georges Cuvier 2. ábra és Charles Bell a fajok közötti különbségekre koncentrált, amelyek megengedik, hogy „létezésük körülményeihez” (conditions of existence) alkalmazkodjanak. Szerintük a Teremtő előrelátása alkotta meg pl. a különböző végtagokat ehhez
2.) Étienne Geoffroy Saint-Hilaire 2. ábra és Owen: „unity of type”- az élőlények közötti hasonlóság a lényeg (Owen nevezte el őket homológiáknak): a végtagok is egyetlen alapterv módosulásai- a Teremtő egy terv alapján teremtette ezeket az élőlényeket- az alkalmazkodások másodlagosak!

A fenti kutatóknál kicsit később, a XIX.század huszas éveiben kutatott a nagy balti német tudós Carl Ernst von Baer (1791-1876). Őt tekintik az összehasonlító embriológia atyjának (nem egyedül csak Őt, többek között Arisztotelész is végzett összehasonlító embriológiai vizsgálatokat). Felfedezései közé tartoznak a gerinchúr, az emlős- és az emberi „pete” (petesejt). Nagyszámú embrió összehasonlítása után felfigyelt rá, hogy a gerincesek korai embrióinak közös sajátságai vannak.
Állításait ma is aligha lehetne jobban megfogalmazni; von Baer törvényeiként máig minden evolúcióról és fejlődésbiológiáról szóló könyv alapvető állításai.
1. Azok a sajátságok, amelyek minden gerincesre jellemzőek, mint a háti oldalon elhelyezkedő agy és gerincvelő, a gerinchúr jelenléte, a szomiták, az aortaívek, a zsigeri (pharyngealis) ívek korábban láthatók a fejlődés folyamán, mint azok a sajátságok, amelyek a különböző gerinces osztályok képviselői között tesznek különbséget (pl. a négylábúak lábai, a madarak tollai, az emlősök szőre). Így egy korai fejlődési stádiumban, a gasztrulációt és neurulációt követően, a halak, kétéltűek, hüllők, madarak és emlősök embriói azonos külsejűek – és belsejűek – de ahogy a fejlődés előre halad, fejlődési útjuk elágazik és felismerhető, melyik osztályba tartozó embrióval állunk szemben, majd a besorolás az idő előrehaladtával egyre pontosabb lehet: eldönthető később, mely rend, család, faj képviselője a vizsgálat tárgya .
2. A kevésbé általános jellegek az általánosabbakból fejlődnek ki, mindaddig, amíg a legspeciálisabbak is megjelennek: Minden gerinces embrión először bőr jelenik meg, majd csak később a specifikus képletek. A végtagok is eleinte hasonlóak
3.) egy adott faj embriója ahelyett, hogy az „alacsonyabbrendű” (ti. az élet történetében előbb megjelenő) fajok felnőtt állapotain mennének át, egyre inkább eltérnek tőle: Pl az embrionális gerincesek kopoltyúszerű képletei nem hasonlítanak a felnőtt halakéira, hanem az embrionális halakéira/gerincesekéire
4.) a „magasabbrendű” állat embriója sohasem hasonlít egy „alacsonyabbrendű” felnőtt állatra, hanem csak annak embriójára
Megfigyeléseit így összegezte:
„Az általánosabb sajátságok, amelyek közösek egy állatcsoport minden tagjánál korábban fejlődtek ki, mint azok a speciálisabb sajátságok, amelyek a csoport különböző tagjait megkülönböztetik.”
Felhívta a figyelmet arra is, hogy az összes gerinces fejlődése azonos mintázat szerint zajlik: a három csíralemezből ugyanazok a szervek fejlődnek ki minden csoportnál
A korai közös szakaszt 1983 óta Sander nyomán filotipikus stádiumnak (phylotypic stage) nevezzük. A filotipikus stádium megjelenése különösen izgalmas tény annak tudatában, hogy az embrió fejlődés bevezető stádiumai – hasadás, gasztruláció – mennyire eltérnek a különféle gerinces osztályokban.
Még az osztályokon belül is – pl. emlősök – a korai stádiumok eltérőek lehetnek.
Von Baer 3. ábra törvénye tehát csakis akkor érvényes, ha a filotipikus stádium a kiindulási pont. Úgy látszik a filotipikus stádium egyfajta szűk keresztmetszet (szokásos a szűk keresztmetszetet bottleneck = üvegnyak névvel illetni; sokféle egyéb értelemben is használják, pl. a populációs genetikában), amelyen minden gerinces embriónak át kell mennie.

Mint már említettük, a kezdeti különbségek után minden gerinces fejlődése egy alapformába torkollik 4. ábra, amelynek jellemzői a velőcső, a gerinchúr (notochord), a szomiták, az oldalsó mezodermalemezek, az előbél a kopoltyúzsebekkel és ívekkel, és egy hasi elhelyezkedésű szívvel. A gerinceseknél a gerincoszlop a gerinchúr köré épül fel.
9.4. ábra - Általános gerinces filotipikus stádium vagy alaptestterv. Ebben a szakaszban a gerincesek alapvető jellegei kimunkáltak: háti helyzetű velőcső, gerinchúr, előbél a kopoltyúnyílásokkal (hasi helyzetű szívvel, amely nincsen az ábrán), szemhólyagokkal és szomitákkal
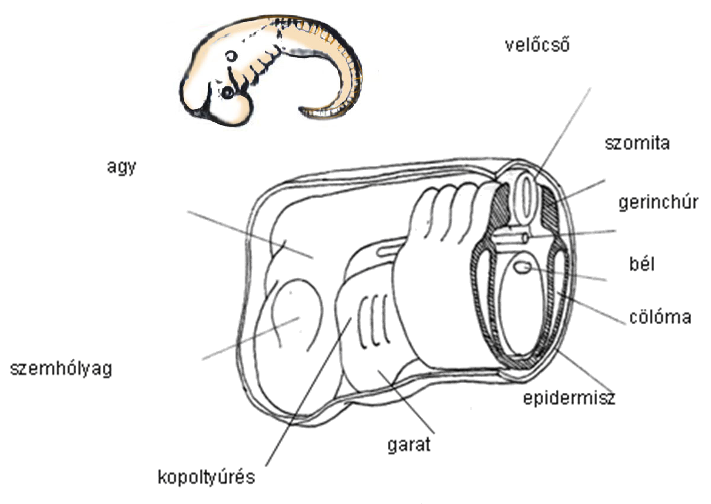 |
A filotipikus stádiumban az idegrendszer egy zárt cső formájában van jelen, amely kissé kitágul elől, ahol az agy fejlődik majd.
Az agy részei megkezdik differenciálódásukat: a velőcsövön megvastagodások és szűkületek (konstrikciók) jelennek meg, és a szemhólyagok is megjelennek az előagy-középagy határán.
Az epidermális epitélium lapszerű megvastagodásokat, ún. plakodokat alkot. A plakodok befűződnek, és szemlencsét és egyéb érzékszerveket alakítanak ki: a nazális plakodok szaglógödrökké alakulnak, az otikus plakodok otikus vezikulumokká fejlődnek, amelyből a belső fül labirintusa és a csiga fejlődik.
A gerinchúr két oldalán a mezoderma szomitákra oszlik fel.
Az előbél alatt a csőszerű szívkezdemények fúzionálnak és kialakítják a hasi elhelyezkedésű szívet.
A legkülönösebb a garati (pharyngealis) régió fejlődése. Evolúciójának örök jeleként a humán embrión is megvannak azok az előbéli kitüremkedések, amelyeket garati (zsigeri) „kopoltyú zsebekként” ismerünk. Szoros kapcsolatban állnak a „kopoltyú” artériákkal, a porcos „kopoltyú ívekkel” (zsigeri ívek) és az elsődleges állkapoccsal, amely a porcos első zsigeri ívből származik. 5. ábra
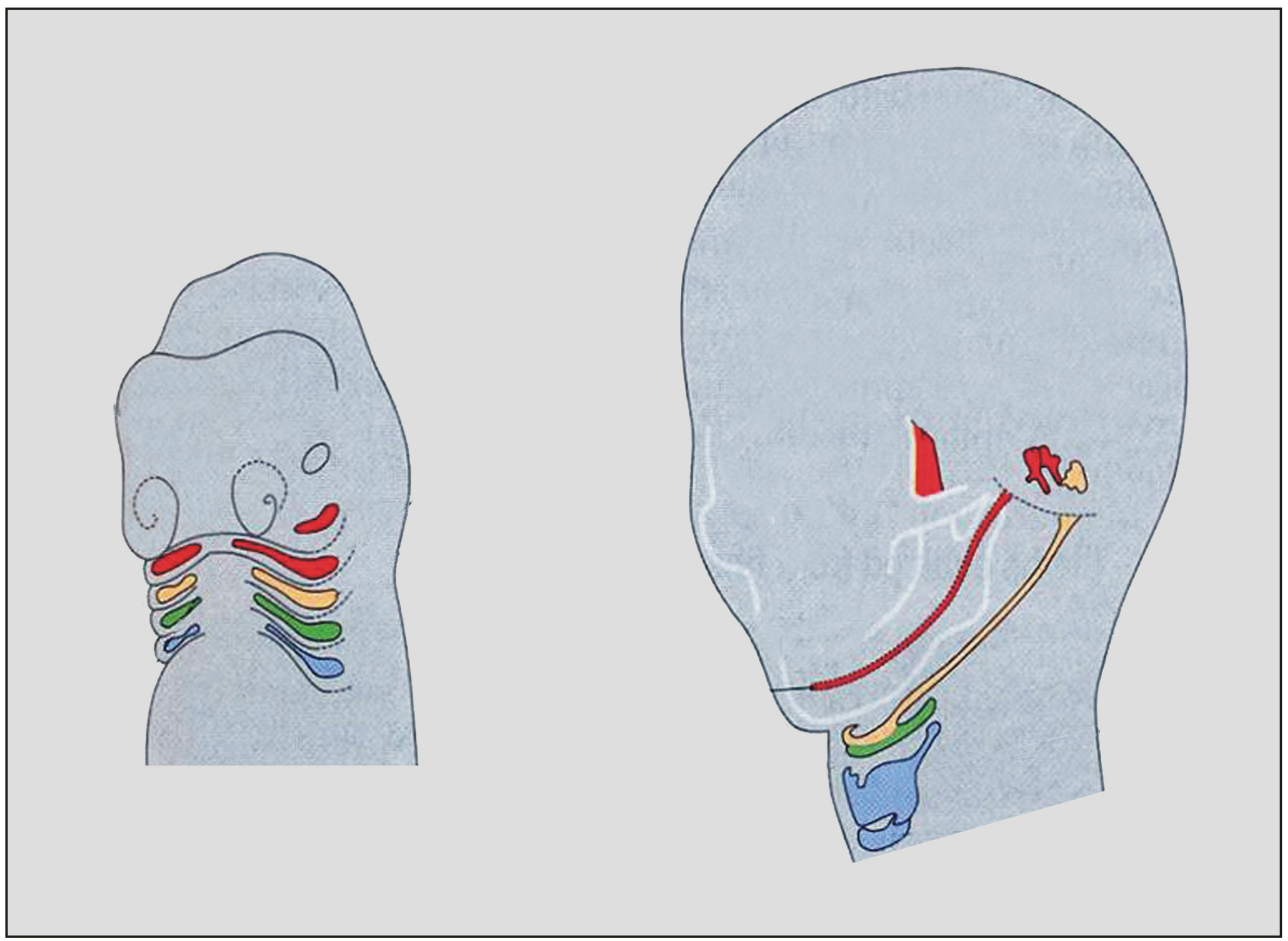
Az evolúció egyik alapkérdése, hogy a korábbi és későbbi fejlődési periódusok nagyfokú különbözősége dacára miért alakul ki a filotipikus stádium? 6. ábra
Csak mostanában született valószínű válasz a kérdésre: az átmeneti (tranzicionális) struktúrákra szükség van a további fejlődés megszervezéséhez. Pl. a gerinchúr és annak előd (prekurzor) sejtjei – az ősbél tetejének középvonalban elhelyezkedő sejtjei – jeleket bocsájtanak ki, amelyek a gerincvelő alaplemezének kialakulását indukálják és iniciálják a szklerotom leválását a szomitákról és a szklerotom sejtjeit céljuk felé vonzzák. A gerinchúrból kiinduló jelek tehát instruálják a szklerotom sejtjeit a csigolyák létrehozására a gerinchúr körül.
A tranzicionális struktúrák szervező szerepét elfogadva is megmarad a kérdés: miért
nem haladhat a fejlődés más úton?
Logikus, hogy egy olyan bonyolult vállalkozás, mint a fejlődés megszervezése nehezen változtatható és lerövidíthető. Azok a megoldások, amelyeket az evolúció évmilliók alatt megtalált, nehezen változtathatók és helyettesíthetők. Semmiképpen nem ősi felnőtt állapotok egymásra következésének vagyunk tehát tanúi, hanem filogenetikailag ősi fejlődési útvonalak – nem feltétlenül gazdaságos – megismétlésének.
Filotipikus stádium más törzseknél
A gerincesek törzse nem egyedi abban, hogy képviselőinek fejlődésében tükröződik az evolúció. A spirális osztódás számos ősszájú törzs (Annelida, Mollusca) esetén megfigyelhetők. A fejlődési útvonalak, amelyek spirális osztódással kezdődnek, gyakran trochophora-szerű lárván át haladnak.7. ábra
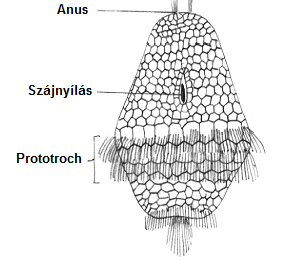
Az onto- és filogenezis szoros kapcsolatban van, ami nem meglepő, hiszen a filogenezis nem más, mint lassan módosuló ontogenezisek megszakítatlan sora.